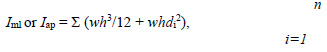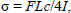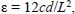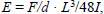Biewener A.A. (1991) Musculoskeletal design in relation to body size. Journal of Biomechanics 24 Suppl 1, 19-29. |
Biewener A.A., Bertram J.E. (1994) Structural response of growing bone to exercise and disuse. Journal of Applied Physiology 76, 946-955. |
Bikle D.D., Halloran B.P. (1999) The response of bone to unloading. Journal of Bone and Mineral Metabolism 17, 233-244. |
Boudreaux R.D., Swift J.M., Gasier H.G., Wiggs M.P., Hogan H.A., Fluckey J.D., Bloomfield S.A. (2014) Increased resistance during jump exercise does not enhance cortical bone formation. Medicine and Science in Sports and Exercise 46, 982-989. |
Burr D.B., Martin R.B., Schaffler M.B., Radin E.L. (1985) Bone remodeling in response to in vivo fatigue microdamage. Journal of Biomechanics 18, 189-200. |
Chen M.M., Yeh J.K., Aloia J.F., Tierney J.M., Sprintz S. (1994) Effect of treadmill exercise on tibial cortical bone in aged female rats: a histomorphometry and dual energy x-ray absorptiometry study. Bone 15, 313-319. |
Clarke B (2008) Normal bone anatomy and physiology. Clinical Journal of the American Society of Nephrology 3, S131-139. |
Edwards W.B., Ward E.D., Meardon S.A., Derrick T.R. (2009) The use of external transducers for estimating bone strain at the distal tibia during impact activity. Journal of Biomechanical Engineering 131, 051009. |
Forwood M.R. (1996) Inducible cyclo-oxygenase (COX-2) mediates the induction of bone formation by mechanical loading in vivo. Journal of Bone and Mineral Research 11, 1688-1693. |
Frost H.M. (2003) Bone’s mechanostat: a 2003 update. The Anatomcial Record. Part A Discoveries in Molecular Cellular and Evolutionary Biology 275, 1081-101. |
Fuchs R.K., Bauer J.J., Snow C.M. (2001) Jumping improves hip and lumbar spine bone mass in prepubescent children: a randomized controlled trial. Journal of Bone and Mineral Research 16, 148-156. |
Garnero P., Borel O., Gineyts E., Duboeuf F., Solberg H., Bouxsein M.L., Christiansen C., Delmas P.D. (2006) Extracellular post-translational modifications of collagen are major determinants of biomechanical properties of fetal bovine cortical bone. Bone 38, 300-309. |
Hattner R., Epker B.N., Frost H.M. (1965) Suggested sequential mode of control of changes in cell behaviour in adult bone remodelling. Nature 206, 489-490. |
Honda A., Umemura Y., Nagasawa S. (2001) Effect of high-impact and low-repetition training on bones in ovariectomized rats. Journal of Bone and Mineral Research 16, 1688-1693. |
Hsieh Y.F., Silva M.J. (2002) In vivo fatigue loading of the rat ulna induces both bone formation and resorption and leads to time-related changes in bone mechanical properties and density. Journal of Orthopedic Research 20, 764-771. |
Huang T.H., Lin S.C., Chang F.L., Hsieh S.S., Liu S.H., Yang R.S. (2003) Effects of different exercise modes on mineralization, structure, and biomechanical properties of growing bone. Journal of Applied Physiology 95, 300-307. |
Johannsen N., Binkley T., Englert V., Neiderauer G., Specker B. (2003) Bone reponse to jumping is site-specific in children: a randomized trial. Bone 33, 533-539. |
Ju Y.I., Sone T., Okamoto T., Fukunaga M. (2008) Jump exercise during remobilization restores integrity of the trabecular architecture after tail suspension in young rats. Journal of Applied Physiology 104, 1594-1600. |
Judex S., Carlson K.J. (2009) Is bone’s response to mechanical signals dominated by gravitational loading?. Medicine and Science in Sports and Exercise 41, 2037-2043. |
Kato T., Terashima T., Yamashita T., Hatanaka Y., Honda A., Umemura Y. (2006) Effect of low-repetition jump training on bone mineral density in young women. Journal of Applied Physiology 100, 839-843. |
Klein-Nulend J., van Oers R.F., Bakker A.D., Bacabac R.G. (2014) Nitric oxide signaling in mechanical adaptation of bone. Osteoporosis International 25, 1427-1437. |
Lin H.S., Huang T.H., Mao S.W., Tai Y.S., Chiu H.T., Cheng K.Y.B., Yang R.S. (2011) A Short-Term Free-Fall Landing Enhances Bone Formation and Bone Material Properties. Journal of Mechanics in Medicine and Biology 11, 1125-1139. |
Lin H.S., Huang T.H., Wang H.S., Mao S.W., Tai Y.S., Chiu H.T., Cheng K.Y., Yang R.S. (2013) Short-term free-fall landing causes reduced bone size and bending energy in femora of growing rats. Journal of Sports Science and Medicine 12, 1-9. |
Notomi T., Lee S.J., Okimoto N., Okazaki Y., Takamoto T., Nakamura T., Suzuki M. (2000) Effects of resistance exercise training on mass, strength, and turnover of bone in growing rats. European of Journal Applied Physiology 82, 268-274. |
Notomi T., Okazaki Y., Okimoto N., Tanaka Y., Nakamura T., Suzuki M. (2002) Effects of tower climbing exercise on bone mass, strength, and turnover in orchidectomized growing rats. Journal of Applied Physiology 93, 1152-1158. |
Notomi T., Okimoto N., Okazaki Y., Tanaka Y., Nakamura T., Suzuki M. (2001) Effects of tower climbing exercise on bone mass, strength, and turnover in growing rats. Journal of Bone and Mineral Research 16, 166-174. |
Parfitt A.M., Drezner M.K., Glorieux F.H., Kanis J.A., Malluche H., Meunier P.J., Ott S.M., Recker R.R. (1987) Bone histomorphometry: standardization of nomenclature, symbols, and units. Report of the ASBMR Histomorphometry Nomenclature Committee. Journal of Bone and Mineral Research 2, 595-610. |
Petit M.A., McKay H.A., MacKelvie K.J., Heinonen A., Khan K.M., Beck T.J. (2002) A randomized school-based jumping intervention confers site and maturity-specific benefits on bone structural properties in girls: a hip structural analysis study. Journal of Bone and Mineral Research 17, 363-372. |
Robling A.G. (2009) Is bone’s response to mechanical signals dominated by muscle forces?. Medicine and Science in Sports Exercise 41, 2044-2049. |
Robling A.G., Castillo A.B., Turner C.H. (2006) Biomechanical and molecular regulation of bone remodeling. Annual Review of Biomedical Engineering 8, 455-498. |
Robling A.G., Niziolek P.J., Baldridge L.A., Condon K.W., Allen M.R., Alam I., Mantila S.M., Gluhak-Heinrich J., Bellido T.M., Harris S.E., Turner C.H. (2008) Mechanical stimulation of bone in vivo reduces osteocyte expression of Sost/sclerostin. Journal of Biologcial Chemistry 283, 5866-5875. |
Rubin C.T., Lanyon L.E. (1985) Regulation of bone mass by mechanical strain magnitude. Calcified. Tissue International 37, 411-417. |
Schulte F.A., Zwahlen A., Lambers F.M., Kuhn G., Ruffoni D., Betts D., Webster D.J., Muller R. (2013) Strain-adaptive in silico modeling of bone adaptation--a computer simulation validated by in vivo micro-computed tomography data. Bone 52, 485-492. |
Sengupta P (2013) The Laboratory Rat: Relating Its Age With Human’s. International Journal of Preventive Medicine 4, 624-630. |
Takata S., Yonezu H., Shibata A., Enishi T., Sato N., Takahashi M., Nakao S., Komatsu K., Yasui N. (2011) Mineral to matrix ratio determines biomaterial and biomechanical properties of rat femur--application of Fourier transform infrared spectroscopy. Journal of Medical Investigation 58, 197-202. |
Van Tran P.T., Vignery A., Baron R. (1982) Cellular kinetics of the bone remodeling sequence in the rat. The Anatomical Record 202, 445-451. |
Turner C.H. (2006) Bone strength: current concepts. Annals of the New York Academy Sciences 1068, 429-446. |
Turner C.H., Burr D.B. (1993) Basic biomechanical measurements of bone: a tutorial. Bone 14, 595-608. |
Umemura Y., Ishiko T., Yamauchi T., Kurono M., Mashiko S. (1997) Five jumps per day increase bone mass and breaking force in rats. Journal of Bone and Mineral Research 12, 1480-1485. |
Umemura Y., Nagasawa S., Honda A., Singh R. (2008) High-impact exercise frequency per week or day for osteogenic response in rats. Journal of Bone and Mineral Metabolism 26, 456-460. |
Verborgt O., Gibson G.J., Schaffler M.B. (2000) Loss of osteocyte integrity in association with microdamage and bone remodeling after fatigue in vivo. Journal of Bone and Mineral Research 15, 60-67. |
Weinreb M., Rodan G.A., Thompson D.D. (1991) Depression of osteoblastic activity in immobilized limbs of suckling rats. Journal of Bone and Mineral Research 6, 725-731. |
Welch J.M., Turner C.H., Devareddy L., Arjmandi B.H., Weaver C.M. (2008) High impact exercise is more beneficial than dietary calcium for building bone strength in the growing rat skeleton. Bone 42, 660-668. |
Welch J.M., Wade J.A., Hillberry B.M., Weaver C.M. (2009) Force platform for rats measures fore and hind forces concurrently. Journal of Biomechanics 42, 2734-2738. |
Welch J.M., Weaver C.M., Turner C.H. (2004) Adaptations to free-fall impact are different in the shafts and bone ends of rat forelimbs. Journal of Applied Physiology 97, 1859-65. |
Xiong J., O’Brien C.A. (2012) Osteocyte RANKL: new insights into the control of bone remodeling. Journal of Bone and Mineral Research 27, 499-505. |
|