The aim of the present research was to propose a new index to describe running coordination and to examine how changes in running velocity would affect this new index. The results confirmed that, when calculated by notational analysis, the % of coordination can illustrate changes in movement organisation, as coordinative patterns tended to be in-phase when athletes approached at high range velocities. The notational analysis of the running coordination showed high levels of intra- and inter-operator reliability, even if the frame rates employed were greater than recommended for running analysis (Payton and Bartlett, 2008). This had been previously observed in expert operators who coded the key events of swimming videos subjectively and obtained high accuracy in comparison to manual digitising of data (Seifert et al., 2006). When the same operator repeatedly identified the key events during the running cycle (except one event), the standard deviation values were lower than the duration of one video frame filmed at 240 Hz (4 ms). This also occurred with the systematic differences between the analyses of independent operators: lower than 4 ms for all key events. Therefore, the uncertainty assumed for the identification of the key events during the running cycle was much less than the time gaps that determine different patterns of coordination (±8% of the stride cycle duration). Greater differences were observed in one specific event (maximum knee extension of the driving leg during the late swing phase), with a standard deviation of 7 ms within and 3 ms between operators. This would suggest that specific training should be performed by operators coding this event and that a reliability test should be conducted to confirm the analysis (Knudson, 1999). Athletes increased relatively more the stride length (15.9%) than the stride frequency (4.9%), as previously suggested (Schache et al., 2014), when augmenting velocity from an easy (4:00 min/km) to a 5000 m pace (3:17 min/km). On the other hand, at the highest velocity, the stride frequency augmented considerably more (19.2%) to assist in the velocity transition to sprinting (Hunter et al., 2004). Running velocities at severe and extreme intensities were similar to those previously reported in national/international level athletes (Bushnell and Hunter, 2007), with athletes surpassing 7 m/s at sprint which has been described as a threshold velocity where torques on the lower limbs increase dramatically (Dorn et al., 2012; Schache et al., 2015). Changes in the running velocity were accompanied by modifications in the DF. As previously reported (Patoz et al., 2022), contact times decreased with increasing velocity until attaining a similar proportion to the flight times, indicating that the work contribution by the contractile muscle was progressively substituted by elastic storage and return by tendons. DF values ranged from 31.3% to 25.6%, which could be categorised as a DFhigh group (Lussiana et al., 2019). Within the stance phase, the brake and push times were reduced with increasing velocity, as previously reported (Lussiana et al., 2019; Patoz et al., 2020), but the proportion of the breaking to pushing phases was approximately one- and two-thirds, respectively. This distribution did not change across the moderate, severe and extreme intensities, showing relatively symmetrical patterns of ground force application across all speeds, which is in contrast with previous results (Cavagna, 2006). Nevertheless, this could be explained by the high DF values of elite athletes in the present study who would minimise vertical displacement of their centre of mass and would favour forward displacement by maintaining greater push to brake times (Lussiana et al., 2019; Patoz et al., 2020). Patterns of coordination seemed to be dependent on the running velocity an also on the stride phase (Figure 4), as athletes tended to decrease time gaps between the observed key events when increasing running velocity from easy pace to sprinting. At touchdown, time gaps between foot contact and the instant the knees were aligned in the running direction represented an average of 8.3% of the running cycle time when sprinting. Considering the stride cycle times of athletes in the present study lasted ≈ 550 ms, time gaps would represent about 45 ms (approximately ten frames of video at 240 Hz) between events. This was similar to what was observed at mid-stance, where time gaps of around 8.9% of the stride cycle time were detected between the hip being on top of the support ankle and the recovery heel being aligned with the support knee. In both cases, shorter time gaps would be expected to occur when sprinting, with the aim of decreasing the breaking forces of the stance and to achieve more horizontal ground reaction force (Kivi et al., 2002), as this would assist in achieving faster velocities (Kakehata et al., 2021). However, the middle-distance runners in the present study may not be familiarised with the required technical modifications related to the change in touchdown geometry (Bushnell and Hunter, 2007) which would include decreasing the knee extension at take-off (Mann and Herman, 1985), increasing the hip angular velocity (Kivi et al., 2002) and the knee flexion at recovery (Williams and Cavanagh, 1987) or modifying the type of foot-strike (Bushnell and Hunter, 2007). Previous research has detected differences between sprinters and distance runners in all these kinematic parameters (Bushnell and Hunter, 2007; Kakehata et al., 2021). For the flight phase coordination, lower time gaps of 1.9% (≈ 10.5 ms or 2–3 frames at 240 Hz) of the difference between the shank being horizontal in the early swing and the maximum knee extension of driving leg were detected. The longer flight-to-contact times may give the athletes enough time to control the position of their moving limbs before touchdown (van Oeveren et al., 2021). Changes in the pattern of coordination with velocity also depended on the stride phase. In the touchdown and mid-stance phases, changes in the % of coordination were lower when transiting at 5000 m velocity than at sprinting. As confirmed by the stride length data, athletes relied on lengthening their stride when increasing the velocity (Table 2). This has been related to very little changes in the overall running form, although some adjustments may occur in the body positioning (Bushnell and Hunter, 2007). However, when sprinting, the observed reduction of the coupling time gaps was probably a consequence of athletes trying to substantially increase the stride frequency (Dorn et al., 2012; Schache et al., 2014), with all the aforementioned technical modifications involved. Conversely, changes in the % of coordination during the flight phase were lower when sprinting. This could be explained by the longer flight times of the 5000 m pace compared to the easy velocity, which would allow athletes to have more time to adjust the inter-limb movements during the flight (van Oeveren et al., 2021). Previous studies have reported that the flight phase remains relatively unaffected by modifications in the task constraints compared to the stance phase (De Wit et al., 2000), which could explain why the main coordinative modifications for sprinting would occur at the stance couplings. Interestingly, no gender differences were observed in the % stride phases nor the patterns of coordination of middle-distance athletes. As expected, male athletes present longer stride lengths and running velocities than females for a certain intensity domain. This has been explained by male athletes being generally taller than females and presenting greater hip flexion during stance, but also by males exerting a greater peak force and peak power than females (Williams et al., 1987). However, these gender differences, as well as others such as the range of motion in the frontal plane and the internal-external rotation of the lower limbs (Takabayashi et al., 2017), did not represent specific patterns of coordination by gender. This could be supported by the lack of gender differences in the % flight-to-contact ratio, contrary to what was previously observed when comparing absolute velocities, where females demonstrated a faster time to peak force and a lower proportion of the stride time in contact with the ground (Barnes et al., 2014). The fact that the female athletes in the present study were part of an elite training group and probably presented a high level of lower-limb strength and/or neuromuscular control could explain similar coordination patterns observed as for male athletes. Also, the fact that gender comparisons were made at similar relative domain intensities and not with absolute velocities would reject the notion of female athletes presenting greater stride frequencies and being less economical than males (Barnes et al., 2014). The present findings confirmed the utility of the proposed notational analysis as a simple and reliable tool to evaluate running coordination in different velocities. The normalised time gaps between key events in the touchdown, mid-stance and flight phases seemed to provide a deep understanding of how athletes self-organise according to the running constraints. Patterns of coordination in competitive middle-distance runners did not completely adhere to Piasenta’s model (Piasenta, 1994), although the increase in the running velocity was related to smaller time gaps between selected events in the running cycle. All evaluation procedures were obtained from a temporal analysis, where no calibration procedures for spatial coordinates’ reconstruction were required. This would ease the application of the coordinative parameters by coaches and practitioners in their training routines to increase the understanding of the athletes’ movements. Further research where the inter-segment couplings could be compared under different constraints (task, organismic or environmental) while running could increase the understanding of how athletes organise their movements. |

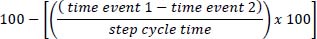

 Barnes K.R., McGuigan M.R., Kilding A.E. (2014) Lower-body determinants of running economy in male and female distance runners. Journal of strength and conditioning research 28, 1289-1297.
Barnes K.R., McGuigan M.R., Kilding A.E. (2014) Lower-body determinants of running economy in male and female distance runners. Journal of strength and conditioning research 28, 1289-1297.

