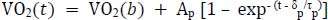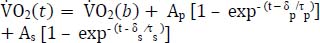Alghannam A.F., Jedrzejewski D., Tweddle M., Gribble H., Bilzon J.L., Betts J.A. (2016) Reliability of Time to Exhaustion Treadmill Running as a Measure of Human Endurance Capacity. International Journal of Sports Medicine 37, 219-223. |
Amann M., Eldridge M.W., Lovering A.T., Stickland M.K., Pegelow D.F., Dempsey J.A. (2006) Arterial oxygenation influences central motor output and exercise performance via effects on peripheral locomotor muscle fatigue in humans. The Journal of Physiology 575, 937-952. |
Amann M., Romer L.M., Subudhi A.W., Pegelow D.F., Dempsey J.A. (2007) Severity of arterial hypoxaemia affects the relative contributions of peripheral muscle fatigue to exercise performance in healthy humans. The Journal of Physiology 581, 389-403. |
Bailey T.G., Birk G.K., Cable N.T., Atkinson G., Green D.J., Jones H., Thijssen D.H. (2012a) Remote ischemic preconditioning prevents reduction in brachial artery flow-mediated dilation after strenuous exercise. American Journal of Physiology Heart and Circulatory Physiology 303, 533-538. |
Bailey T.G., Jones H., Gregson W., Atkinson G., Cable N.T., Thijssen D.H. (2012b) Effect of ischemic preconditioning on lactate accumulation and running performance. Medicine Science in Sports Exercise 44, 2084-2089. |
Barbosa T.C., Machado A.C., Braz I.D., Fernandes I.A., Vianna L.C., Nobrega A.C., Silva B.M. (2015) Remote ischemic preconditioning delays fatigue development during handgrip exercise. Scandinavian Journal of Medicine Science in Sports 25, 356-364. |
Bellini D., Chapman C., Peden D., Hoekstra S.P., Ferguson R.A., Leicht C.A. (2023) Ischaemic preconditioning improves upper-body endurance performance without altering V̇O(2) kinetics. European Journal of Sport Science 23, 1538-1546. |
Bennett S., Brocherie F., Phelan M.M., Tiollier E., Guibert E., Morales-Artacho A.J., Lalire P., Morton J.P., Louis J.B., Owens D.J. (2023) Acute heat stress amplifies exercise-induced metabolomic perturbations and reveals variation in circulating amino acids in endurance-trained males. Experimental Physiology 108, 838-851. |
Bruce R.A., Kusumi F., Hosmer D. (1973) Maximal oxygen intake and nomographic assessment of functional aerobic impairment in cardiovascular disease. American Heart Journal 85, 546-562. |
Byrne C., Lim C.L. (2007) The ingestible telemetric body core temperature sensor: a review of validity and exercise applications. British Journal of Sports Medicine 41, 126-133. |
Casa D.J., Armstrong L.E., Hillman S.K., Montain S.J., Reiff R.V., Rich B.S., Roberts W.O., Stone J.A. (2000) National athletic trainers' association position statement: fluid replacement for athletes. Journal of Athletic Training 35, 212-224. |
Cerezuela-Espejo V., Courel-Ibanez J., Moran-Navarro R., Martinez-Cava A., Pallares J.G. (2018) The Relationship Between Lactate and Ventilatory Thresholds in Runners: Validity and Reliability of Exercise Test Performance Parameters. Frontiers in Physiology 9, 1320. |
Cheung C.H.Y., Khaw M.L., Tam V.C.W., Ying M.T.C., Lee S.W.Y. (2020) Performance evaluation of a portable bioimpedance cardiac output monitor for measuring hemodynamic changes in athletes during a head-up tilt test. Journal of Applied Physiology (1985) 128, 1146-1152. |
Cheuvront S.N., Kenefick R.W., Montain S.J., Sawka M.N. (2010) Mechanisms of aerobic performance impairment with heat stress and dehydration. Journal of Applied Physiology (1985) 109, 1989-1995. |
Cocking S., Landman T., Benson M., Lord R., Jones H., Gaze D., Thijssen D.H.J., George K. (2017) The impact of remote ischemic preconditioning on cardiac biomarker and functional response to endurance exercise. Scandinavian Journal of Medicine Science in Sports 27, 1061-1069. |
Crisafulli A., Tangianu F., Tocco F., Concu A., Mameli O., Mulliri G., Caria M.A. (2011) Ischemic preconditioning of the muscle improves maximal exercise performance but not maximal oxygen uptake in humans. Journal of Applied Physiology (1985) 111, 530-526. |
Cruz R.S., de Aguiar R.A., Turnes T., Pereira K.L., Caputo F. (2015) Effects of ischemic preconditioning on maximal constant-load cycling performance. Journal of Applied Physiology (1985) 119, 961-967. |
da Mota G.R., Willis S.J., Sobral N.D.S., Borrani F., Billaut F., Millet G.P. (2019) Ischemic Preconditioning Maintains Performance on Two 5-km Time Trials in Hypoxia. Medicine Science in Sports Exercise 51, 2309-2317. |
de Groot P.C., Thijssen D.H., Sanchez M., Ellenkamp R., Hopman M.T. (2010) Ischemic preconditioning improves maximal performance in humans. European Journal of Applied Physiology 108, 141-146. |
Dimenna F.J., Fulford J., Bailey S.J., Vanhatalo A., Wilkerson D.P., Jones A.M. (2010) Influence of priming exercise on muscle [PCr] and pulmonary O2 uptake dynamics during 'work-to-work' knee-extension exercise. Respiratory Physiology Neurobiology 172, 15-23. |
Domitrovich J.W., Cuddy J.S., Ruby B.C. (2010) Core-temperature sensor ingestion timing and measurement variability. Journal of Athletic Training 45, 594-600. |
Driver S., Reynolds M., Brown K., Vingren J.L., Hill D.W., Bennett M., Gilliland T., McShan E., Callender L., Reynolds E., Borunda N., Mosolf J., Cates C., Jones A. (2022) Effects of wearing a cloth face mask on performance, physiological and perceptual responses during a graded treadmill running exercise test. British Journal of Sports Medicine 56, 107-113. |
Ely B.R., Cheuvront S.N., Kenefick R.W., Sawka M.N. (2010) Aerobic performance is degraded, despite modest hyperthermia, in hot environments. Medicine Science in Sports Exercise 42, 135-141. |
Enko K., Nakamura K., Yunoki K., Miyoshi T., Akagi S., Yoshida M., Toh N., Sangawa M., Nishii N., Nagase S., Kohno K., Morita H., Kusano K.F., Ito H. (2011) Intermittent arm ischemia induces vasodilatation of the contralateral upper limb. The Journal of Physiological Sciences 61, 507-513. |
Feldmann A.M., Erlacher D., Pfister S., Lehmann R. (2020) Muscle oxygen dynamics in elite climbers during finger-hang tests at varying intensities. Scientific Reports 10, 3040. |
Gao X., Wang A., Fan J., Zhang T., Li C., Yue T., Hurr C. (2024) The effect of ischemic preconditioning on repeated sprint cycling performance: a randomized crossover study. The Journal of Sports Medicine and Physical Fitness. |
Gardner R.N., Sabino-Carvalho J.L., Kim J., Vianna L.C., Lang J.A. (2020) Two weeks of remote ischaemic preconditioning alters sympathovagal balance in healthy humans. Experimental Physiology 105, 1500-1506. |
Gibson N., Mahony B., Tracey C., Fawkner S., Murray A. (2015) Effect of ischemic preconditioning on repeated sprint ability in team sport athletes. Journal of Sports Sciences 33, 1182-1188. |
Gonzalez-Alonso J., Crandall C.G., Johnson J.M. (2008) The cardiovascular challenge of exercising in the heat. The Journal of Physiology 586, 45-53. |
Hinds T., McEwan I., Perkes J., Dawson E., Ball D., George K. (2004) Effects of massage on limb and skin blood flow after quadriceps exercise. Medicine Science in Sports Exercise 36, 1308-1313. |
Incognito A.V., Burr J.F., Millar P.J. (2016) The Effects of Ischemic Preconditioning on Human Exercise Performance. Sports Medicine 46, 531-544. |
Incognito A.V., Doherty C.J., Lee J.B., Burns M.J., Millar P.J. (2017) Ischemic preconditioning does not alter muscle sympathetic responses to static handgrip and metaboreflex activation in young healthy men. Physiological Reports 5. |
James C.A., Willmott A.G., Richardson A.J., Watt P.W., Maxwell N.S. (2016) Ischaemic preconditioning does not alter the determinants of endurance running performance in the heat. European Journal of Applied Physiology 116, 1735-1745. |
Jean-St-Michel E., Manlhiot C., Li J., Tropak M., Michelsen M.M., Schmidt M.R., McCrindle B.W., Wells G.D., Redington A.N. (2011) Remote preconditioning improves maximal performance in highly trained athletes. Medicine Science in Sports Exercise 43, 1280-1286. |
Johnson J.M. (2010) Exercise in a hot environment: the skin circulation. Scandinavian Journal of Medicine Science in Sports 20, 29-39. |
Jones A.M., Grassi B., Christensen P.M., Krustrup P., Bangsbo J., Poole D.C. (2011) Slow component of VO2 kinetics: mechanistic bases and practical applications. Medicine Science in Sports Exercise 43, 2046-2062. |
Jones H., Hopkins N., Bailey T.G., Green D.J., Cable N.T., Thijssen D.H. (2014) Seven-day remote ischemic preconditioning improves local and systemic endothelial function and microcirculation in healthy humans. American Journal of Hypertension 27, 918-925. |
Kido K., Suga T., Tanaka D., Honjo T., Homma T., Fujita S., Hamaoka T., Isaka T. (2015) Ischemic preconditioning accelerates muscle deoxygenation dynamics and enhances exercise endurance during the work-to-work test. Physiological Reports 3. |
Kilding A.E., Sequeira G.M., Wood M.R. (2018) Effects of ischemic preconditioning on economy, VO(2) kinetics and cycling performance in endurance athletes. European Journal of Applied Physiology 118, 2541-2549. |
Kim J., Franke W.D., Lang J.A. (2021) Delayed window of improvements in skin microvascular function following a single bout of remote ischaemic preconditioning. Experimental Physiology 106, 1380-1388. |
Kloner R.A., Rezkalla S.H. (2006) Preconditioning, postconditioning and their application to clinical cardiology. Cardiovascular Research 70, 297-307. |
Korzeniewski B. (2018) Regulation of oxidative phosphorylation is different in electrically- and cortically-stimulated skeletal muscle. Plos One 13, e0195620. |
Korzeniewski B. (2023) V̇O(2) (non-)linear increase in ramp-incremental exercise vs. V̇O(2) slow component in constant-power exercise: Underlying mechanisms. Respiratory Physiology Neurobiology 311, 104023. |
Kuntscher M.V., Kastell T., Altmann J., Menke H., Gebhard M.M., Germann G. (2002) Acute remote ischemic preconditioning II: the role of nitric oxide. Microsurgery 22, 227-231. |
Lang J.A., Kim J. (2022) Remote ischaemic preconditioning - translating cardiovascular benefits to humans. The Journal of Physiology 600, 3053-3067. |
Lang J.A., Kim J., Franke W.D., Vianna L.C. (2019) Seven consecutive days of remote ischaemic preconditioning improves cutaneous vasodilatory capacity in young adults. The Journal of Physiology 597, 757-765. |
Laursen P.B., Francis G.T., Abbiss C.R., Newton M.J., Nosaka K. (2007) Reliability of time-to-exhaustion versus time-trial running tests in runners. Medicine Science in Sports Exercise 39, 1374-1379. |
Ley O., Dhindsa M., Sommerlad S.M., Barnes J.N., Devan A.E., Naghavi M., Tanaka H. (2011) Use of temperature alterations to characterize vascular reactivity. Clinical Physiology and Functional Imaging 31, 66-72. |
Lucia A., Hoyos J., Perez M., Chicharro J.L. (2000) Heart rate and performance parameters in elite cyclists: a longitudinal study. Medicine Science in Sports Exercise 32, 1777-1782. |
Marocolo M., da Mota G.R., Simim M.A.M., Appell Coriolano H.J. (2016) Myths and Facts About the Effects of Ischemic Preconditioning on Performance. International Journal of Sports Medicine 37, e7. |
Marocolo M., Hohl R., Arriel R.A., Mota G.R. (2023) Ischemic preconditioning and exercise performance: are the psychophysiological responses underestimated?. European Journal of Applied Physiology 123, 683-693. |
Michaelides A.P., Tousoulis D., Fourlas C.A., Andrikopoulos G.K., Dilaveris P.E., Aggeli K.D., Psomadaki Z.D., Stefanadis C.I. (2006) The aortic distensibility alteration is an index of influence of ischemic preconditioning to myocardial performance. International Journal of Cardiology 113, 76-81. |
Moro L., Pedone C., Mondi A., Nunziata E., Antonelli Incalzi R. (2011) Effect of local and remote ischemic preconditioning on endothelial function in young people and healthy or hypertensive elderly people. Atherosclerosis 219, 750-752. |
Mulliri G., Sainas G., Magnani S., Palazzolo G., Milia N., Orru A., Roberto S., Marongiu E., Milia R., Crisafulli A. (2016) Ischemic preconditioning reduces hemodynamic response during metaboreflex activation. American Journal of Physiology Regulatory Integrative and Comparative Physiology 310, 777-787. |
Murry C.E., Jennings R.B., Reimer K.A. (1986) Preconditioning with ischemia: a delay of lethal cell injury in ischemic myocardium. Circulation 74, 1124-1136. |
Ogawa T., Koike J., Hirano Y. (2022) The effects of cloth face masks on cardiorespiratory responses and VO(2) during maximal incremental running protocol among apparently healthy men. Scientific Reports 12, 22292. |
Otani H., Kaya M., Tamaki A., Watson P., Maughan R.J. (2016) Effects of solar radiation on endurance exercise capacity in a hot environment. European Journal of Applied Physiology 116, 769-779. |
Ozyener F., Rossiter H.B., Ward S.A., Whipp B.J. (2001) Influence of exercise intensity on the on- and off-transient kinetics of pulmonary oxygen uptake in humans. The Journal of Physiology 533, 891-902. |
Paradis-Deschenes P., Joanisse D.R., Billaut F. (2018) Ischemic Preconditioning Improves Time Trial Performance at Moderate Altitude. Medicine Science in Sports Exercise 50, 533-541. |
Paradis-Deschenes P., Lapointe J., Joanisse D.R., Billaut F. (2020) Similar Recovery of Maximal Cycling Performance after Ischemic Preconditioning, Neuromuscular Electrical Stimulation or Active Recovery in Endurance Athletes. Journal of Sports Science and Medicine 19, 761-771. |
Patterson S.D., Bezodis N.E., Glaister M., Pattison J.R. (2015) The Effect of Ischemic Preconditioning on Repeated Sprint Cycling Performance. Medicine Science in Sports Exercise 47, 1652-1658. |
Paull E.J., Van Guilder G.P. (2019) Remote ischemic preconditioning increases accumulated oxygen deficit in middle-distance runners. Journal of Applied Physiology (1985) 126, 1193-1203. |
Pearson S.J., Hussain S.R. (2015) A review on the mechanisms of blood-flow restriction resistance training-induced muscle hypertrophy. Sports Medicine 45, 187-200. |
Pereira H.M., de Lima F.F., Silva B.M., Kohn A.F. (2020) Sex differences in fatigability after ischemic preconditioning of non-exercising limbs. Biology of Sex Differences 11, 59. |
Periard J.D., Caillaud C., Thompson M.W. (2012) The role of aerobic fitness and exercise intensity on endurance performance in uncompensable heat stress conditions. European Journal of Applied Physiology 112, 1989-1999. |
Racinais S., Alonso J.M., Coutts A.J., Flouris A.D., Girard O., Gonzalez-Alonso J., Hausswirth C., Jay O., Lee J.K., Mitchell N., Nassis G.P., Nybo L., Pluim B.M., Roelands B., Sawka M.N., Wingo J., Periard J.D. (2015) Consensus recommendations on training and competing in the heat. British Journal of Sports Medicine 49, 1164-1173. |
Sabino-Carvalho J.L., Lopes T.R., Obeid-Freitas T., Ferreira T.N., Succi J.E., Silva A.C., Silva B.M. (2017) Effect of Ischemic Preconditioning on Endurance Performance Does Not Surpass Placebo. Medicine Science in Sports Exercise 49, 124-132. |
Sabino-Carvalho J.L., Obeid-Freitas T., Paula-Ribeiro M., Lopes T.R., Ferreira T.H.N., Succi J.E., Silva A.C., Silva B.M. (2019) Ischemic preconditioning boosts post-exercise but not resting cardiac vagal control in endurance runners. European Journal of Applied Physiology 119, 621-632. |
Salvador A.F., De Aguiar R.A., Lisboa F.D., Pereira K.L., Cruz R.S., Caputo F. (2016) Ischemic Preconditioning and Exercise Performance: A Systematic Review and Meta-Analysis. International Journal of Sports Physiology and Performance 11, 4-14. |
Sawka M.N., Leon L.R., Montain S.J., Sonna L.A. (2011) Integrated physiological mechanisms of exercise performance, adaptation, and maladaptation to heat stress. Comprehensive Physiology 1, 1883-928. |
Shimizu M., Konstantinov I.E., Kharbanda R.K., Cheung M.H., Redington A.N. (2007) Effects of intermittent lower limb ischaemia on coronary blood flow and coronary resistance in pigs. Acta Physiologica (Oxf) 190, 103-109. |
Stucky F., Vesin J.M., Kayser B., Uva B. (2018) The Effect of Lower-Body Positive Pressure on the Cardiorespiratory Response at Rest and during Submaximal Running Exercise. Frontiers in Physiology 9, 34. |
Teixeira A.L., Gangat A., Bommarito J.C., Burr J.F., Millar P.J. (2023) Ischemic Preconditioning Acutely Improves Functional Sympatholysis during Handgrip Exercise in Healthy Males but not Females. Medicine Science in Sports Exercise 55, 1250-1257. |
Thijssen D.H., Maxwell J., Green D.J., Cable N.T., Jones H. (2016) Repeated ischaemic preconditioning: a novel therapeutic intervention and potential underlying mechanisms. Experimental Physiology 101, 677-692. |
van Beekvelt M.C., Borghuis M.S., van Engelen B.G., Wevers R.A., Colier W.N. (2001) Adipose tissue thickness affects in vivo quantitative near-IR spectroscopy in human skeletal muscle. Clinical Science (Lond) 101, 21-28. |
Van Cutsem J., Roelands B., De Pauw K., Meeusen R., Marcora S. (2019) Subjective thermal strain impairs endurance performance in a temperate environment. Physiology Behavior 202, 36-44. |
Wang A., Brothers R.M., Hurr C. (2023) Application of blood flow restriction in hypoxic environment augments muscle deoxygenation without compromising repeated sprint exercise performance. Experimental Physiology 108, 728-739. |
Wiggins C.C., Constantini K., Paris H.L., Mickleborough T.D., Chapman R.F. (2019) Ischemic Preconditioning, O2 Kinetics, and Performance in Normoxia and Hypoxia. Medicine Science in Sports Exercise 51, 900-911. |
Wilk M., Krzysztofik M., Jarosz J., Krol P., Leznicka K., Zajac A., Stastny P., Bogdanis G.C. (2021) Impact of Ischemic Intra-Conditioning on Power Output and Bar Velocity of the Upper Limbs. Frontiers in Physiology 12, 626915. |
Wilkinson D.M., Carter J.M., Richmond V.L., Blacker S.D., Rayson M.P. (2008) The effect of cool water ingestion on gastrointestinal pill temperature. Medicine Science in Sports Exercise 40, 523-528. |
Williams N., Russell M., Cook C.J., Kilduff L.P. (2021) Effect of Ischemic Preconditioning on Maximal Swimming Performance. The Journal of Strength Conditioning Research 35, 221-226. |
Zhou K., Yang B., Zhou X.M., Tan C.M., Zhao Y., Huang C., Liao X.B., Xiao H.B. (2007) Effects of remote ischemic preconditioning on the flow pattern of the left anterior descending coronary artery in normal subjects. International Journal of Cardiology 122, 250-251. |
|