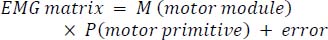|
|
|
| |
| ABSTRACT |
|
Basketball victory relies on an athlete’s skill to make precise shots at different distances. While extensive research has explored the kinematics and dynamics of different shooting distances, the specific neuromuscular control strategies involved remain elusive. This study aimed to compare the differences in muscle synergies during basketball shooting at different distances, offering insights into neuromuscular control strategies and guiding athletes’ training. Ten skilled shooting right-handed male basketball players participated as subjects in this experiment. Electromyographic (EMG) data for full-phase shooting were acquired at short (3.2 m), middle (5.0 m), and long (6.8 m) distances. Non-negative matrix decomposition extracted muscle synergies (motor modules and motor primitives) during shooting. The results of this study show that all three distance shooting can be broken down into three synergies and that there were differences in the synergies between short and long distances, with differences in motor primitive 1 and motor primitive 2 at the phase of 45% - 59% (p < 0.001, t* = 4.418), and 78% - 88% (p < 0.01, t* = 4.579), respectively, and differences in the motor module 3 found in the differences in muscle weights for rectus femoris (RF) (p = 0.001, d = -2.094), and gastrocnemius lateral (GL) (p = 0.001, d = -2.083). Shooting distance doesn't affect the number of muscle synergies in basketball shooting but alters synergy patterns. During long distance shooting training, basketball players should place more emphasis on the timing and synergistic activation of upper and lower limbs, as well as core muscles. |
| Key words:
Scoring, Muscle Synergy, Coordination, Athletes
|
Key
Points
- The present study investigated and compared muscle synergies in basketball shooting at three distances (short, middle, and long) to determine whether different neuromuscular coordination and control strategies exist.
- Kinematic and EMG data were collected from basketball players shooting from distances of 3.2 m, 5.0 m, and 6.8 m. and analyzed using NNMF to identify muscle synergies.
- The neuromuscular control strategies for all shooting distances can be explained by three muscle synergies. The number of muscle synergies remains constant across distances, but their patterns vary with shooting distance.
- The results recommend that novice basketball players emulate the muscle synergy patterns of elite players in their upper limbs, lower limbs, and core to enhance their shooting technique. For players proficient in short distance but not long distance shooting, we advise adjusting the timing and intensity of lower limb activation, and leveraging the energy generated from jumping and body twists to improve long distance shooting performance.
|
Basketball is one of the most popular sports worldwide, and shooting stands as the fundamental skill and primary scoring method (Okazaki et al., 2015). Proficiency in shooting from various distances improves a team’s likelihood of winning a basketball game (Csataljay et al., 2009; Okazaki and Rodacki, 2012). Previous studies investigated external manifestations such as human kinematics (Cabarkapa et al., 2021), ball kinematics (Miller and Bartlett, 1996), and more to understand the shooting mechanism at various distances and guide basketball player training. However, very few studies are available on the inner neuromuscular control strategy of shooting from different distances, and muscle synergy can reflect the neuromuscular control strategy of athletes to complete the movement technique after years of training (Sawers et al., 2015). Scholars suggest that individual muscles are not activated in isolation while orchestrating movement execution inside the human central nervous system. Instead, the central nervous system is proposed to govern a cluster of lower modules, each comprising the weighted interactions of multiple muscles (Safavynia et al., 2011). These modules are organized in a synchronized spatial and temporal manner, collectively contributing to the seamless completion of a given movement (Frère and Hug, 2012). This is referred to as muscle synergy. The decomposition of surface EMG signals from multiple muscles enables the extraction of muscle synergies, representing them as motor modules (weights assigned to each muscle) and motor primitives (degree of activation over time) (Cheung et al., 2009). This method is widely adopted by many scholars to investigate the neuromuscular coordination and control strategies of motor skills (Safavynia et al., 2011). Muscle synergies in the lower limbs of experienced cyclists have been found to exhibit shared neuromuscular control strategies (Hug et al., 2010). Similarly, studies on the muscle synergy in whole-body complex movement skills, it was found that skilled athletes have great similarity in their muscle synergy when completing the same complex technical movement (Frère and Hug, 2012). Furthermore, research indicates that increasing task difficulty or experiencing fatigue does not alter the structure of muscle synergy, but rather modifies the pattern of synergy (Hajiloo et al., 2020). Basketball shooting is also a complex movement skills (Safavynia and Ting, 2012), with task difficulty escalating as the shooting distance increases, thereby altering the shooting movement patterns of skilled basketball player (Okazaki and Rodacki, 2012). However, it is not clear whether different neuromuscular control strategies exist for skilled shooting basketball players to complete shooting tasks at different distances, and if they do, then these differences may be the key to skilled shooting basketball players being able to accurately complete shots at various distances. The purpose of this study was to compare the muscle synergies of skilled shooting basketball players when shooting at different distances, whether different neuromuscular coordination and control strategies exist, and to use these differences to provide recommendations for coaches and athletes training. We hypothesized that skilled shooting basketball players exhibit different muscle synergies when completing shots at varying distances. SubjectsAll subjects were selected from the local youth basketball sports teams, and the 10 right-handed shooting male athletes (age: 18.1 ± 0.7 years; height: 185.8 ± 5.3 cm; weight: 78.8 ± 6.3 kg; years of training: 5.3 ± 1.6 years) with the highest shooting percentages were selected as subjects, which was the same number of subjects selected for a previous study (Nakano et al., 2020) on shooting from different distances. All subjects had not suffered any injuries during the six-month period and were in good health during the experiment. Subjects were advised to avoid excessive exercise or physical activity and to rest well before the experiment for better result accuracy. Before the experiment, and each subject obtained written informed signed consent after a detailed explanation of the testing protocol, the possible risks involved, and the right to terminate participation at will. All procedures were in accordance with the Declaration of Helsinki 1975, as revised in 1996, and were approved by the Jeonbuk University Ethics Committee (JBNU2022-04-008-002).
Experimental ProcedureAll experiments were conducted between 15:00-17:00 each day. Subjects initially underwent a ten-minute standardized warm-up, followed by a three-minute break before the experiment commenced. Each subject executed shots at short (3.2 m), middle (5.0 m), and long (6.8 m) distances (Figure 1) in their preferred manner, ensuring comfort in the technical movement. Adequate rest intervals between shots were taken to prevent fatigue from consecutive attempts.
Collection of dataA whole body position data of each subject was captured using motion capture system (13 infrared cameras; Motive 2.2.0, Natural Point, Inc., Corvallis, OR, USA) at a sampling rate of 120 Hz with 45 reflective skin markers (14mm) on each subject. EMG signals were gathered during basketball shooting through Trigno wireless sensors (Trigno Avanti Sensor, Delsys, Natick, MA, USA). A partial skin preparation involving cleaning and shaving was conducted to reduce contact surface impedance between the skin and electrode (Li et al., 2023). EMG signals from the right flexor carpi radialis brevis (FCR), biceps brachii (BB), triceps brachii (TB), anterior deltoid (AD), latissimus dorsi (LD), external oblique (EOM), erector spinae (ES), rectus femoris (RF), and gastrocnemius lateral (GL) muscles were recorded by placing electrodes longitudinally with respect to the muscle fiber arrangement at a sampling frequency of 1200 Hz. Subsequently, the acquired data from the motion capture system were meticulously modeled in Visual 3D (C-Motion, Inc., Germantown, MD, USA). Model and ball kinematic data were calculated and filtered using a low-pass filter (16 Hz, Butterworth filter, 4th order). The shooting phase of the experiment was outlined based on the ball position and body kinematic data: sinking stage, lifting ball stage, take-off stage, release stage (Figure 2). A delay of 30 ms between muscle activation and movement initiation was selected, according to the shooting phase for capturing the EMG data throughout the entire motion (Norman and Komi, 1979).
Data analysisEMG data from each shooting phase intercepted according to kinematic stages were imported into Matlab R2021a for processing. The EMG signals were band-pass filtered (20–450 Hz, Butterworth filter, 4th order), full-wave rectified, and smoothed with a zero-lag low-pass filter (6 Hz, Butterworth filter, 4th order) to obtain linear envelopes and normalized by time was performed to obtain 100 data points for each shooting action (Barroso et al., 2014). The final EMG data was normalized to the maximum activity level at all shooting distances. In order to demonstrate the reproducibility of the EMG data in this experiment, correlation tests were conducted on the normalized post-EMG data obtained from the three measurements in each shooting test for each subject. Subsequently, for each subject and each shooting distance, two datasets with the highest correlation (where all muscle correlations were greater than 0.9) were selected for synergy extraction (Aoyama et al., 2022). Non-negative matrix factorization (NNMF) was applied to decompose the pre-processed EMG signal matrix to decompose and derive muscle synergies. The algorithmic formulation is as follows (Lee and Seung, 2000):
The EMG matrix is an initial matrix of m rows and n columns (m = 9 is the number of muscles, and n = 100 is the number of normalized time points). M is a matrix comprising m rows and α columns (α is the number of muscle synergies), and P is a matrix comprising α rows and n columns. Error is defined as the difference between the initial and reconstructed EMG matrices. The algorithm is based on iteratively updating the initial random guesses for M and P, converging to a locally optimal matrix decomposition. The algorithm is repeated at least 20 times for each subject to avoid local optimal solutions. The overall optimal solution (i.e., minimizing the squared error between the initial and reconstructed EMG patterns) was retained. Using the algorithm, the pre-processed EMG matrices for each subject’s three distance shots were decomposed into muscle synergy weighted (M) and activated (P) matrices based on temporal order. The term ‘motor module’ refers to the muscle synergy weighted matrix (M), whereas ‘motor primitives’ refers to the activation matrix (P). Each motor module is represented as an (m x α) vector, with its components reflecting the relative contribution (weight) of each muscle to that specific synergy, and the corresponding motor primitives indicate the module’s activation patterns over the time series. There are two common approaches for determining the number of synergies, both requiring the variance accounted for (VAF) to be calculated first. The first approach is to choose the number of synergies that account for more than 0.9 of the VAF (Torres-Oviedo and Ting, 2010),whereas the second approach uses the inflection point of the best linear fit result as the criterion (Clark et al., 2010). The average total VAF was defined as follow :
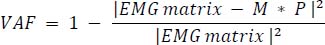 To determine the number of muscle synergies, VAF from 1 to 9 was calculated based on the number of muscles sampled. And, VAF (Figure 3) per distance shot for each subject at each number of synergies was calculated to ensure that the muscle synergies accurately explained the neuromuscular control strategy at each distance shot for each subject. When the motor module was 3, the short distance shooting VAF = 0.948 ± 0 .022, the middle distance shooting VAF = 0.951 ± 0.022, and the long distance shooting VAF = 0.98 ± 0.010. As a result, we limited the number of synergistic elements (α) to 3 and extracted the motor modules and motor primitives for muscle synergism when subjects shot at the three distances. The k-means clustering algorithm was utilized to classify the motor modules retrieved from each distance shot for the ten subjects (Pan et al., 2023). Since VAF indicates that there are 3 motor modules, K was set to 3. Considering that the initial clustering center of mass influences the k-means solution, randomness was reduced by repeating the process 50 times with different initial centers of mass. Profile scores were computed for all clustering outcomes to assess the optimal clustering result. The result with the highest profile score was selected as the optimal clustering outcome, and motor modules with their corresponding motor primitives were outputted. Each distance was clustered using this approach, and the motor modules and motor primitives for all distances were obtained (Figure 4a, Figure 4b and Figure 4c).
Statistical analysisCosine similarity (CS) analysis was used to test the similarity between the motor modules of all subjects at each shooting distance (Hagio and Kouzaki, 2015). A CS value closer to 1 indicates a higher degree of similarity. Based on previous relevant studies (Cheung et al., 2005; Santuz et al., 2017), CS threshold was set at 0.8 in this study. High similarity was defined as CS > 0.8 between motor modules, and paired-sample t-tests were conducted for motor modules with a CS < 0.8 to elucidate which changes in muscle weight led to alterations in motor modules (Park and Caldwell, 2022). Meanwhile, one-dimensional Statistical Parametric Mapping (SPM1d) was used to perform a one-way repeated-measures ANOVA on the motor primitives of all subjects at three shooting distances, analyzing their temporal changes (Pan et al., 2023). Multiple comparisons were performed on different distances of this motor primitive if there was a difference, with the significant level critical threshold set at α < 0.017 (0.05/number of groups). The values of SPM (F) and SPM (t*) were computed during the analysis. The statistics of F and t* serve the same purpose as effect sizes and can be used as indicators of practical significance (Pataky et al., 2015). All statistical analyses were performed in Matlab R2021a.
Characteristics of the motor modules of shooting movements at different distancesThe motor modules of all subjects at different shooting distances are shown in Figure 4a, Figure 4b and Figure 4c, with each shooting distance being composed of three motor modules. These modules have specific dominant muscles corresponding to different shooting phases, i.e., in motor module 3 (M3), the CS of short and long distance shooting is 0.782 (CS < 0.8, Table 1). Therefore, a multiple paired t-test was performed on all muscle weights of this data group. Results showed significant differences (p < 0.01) in BB (p = 0.005, d = 1.446, Figure 5), EOM (p = 0.001, d = -3.710, Figure 5), RF (p = 0.001, d = -2.094, Figure 5), and GL (p = 0.001, d = -2.083, Figure 5) muscle weights at 6.8 m and 3.2 m in motor module 3.
Characteristics of the motor primitives of shooting movements at different distancesA difference at the 42% - 54% phase of motor primitive 1 (the activation level of motor module 1) (p < 0.001, F = 8.877, Figure 6) was found. Multiple comparisons of motor primitive 1 between different distances revealed that there was a difference only in the 45% - 59% phase of motor primitives between short distance shooting and long distance shooting (p < 0.001, t* = 4.418, Figure 6); motor module 1 was activated to a significantly greater extent at the lifting ball stage when performing short distance than at long distance shooting. Differences in motor primitive 2 (activation of motor module 2) were seen at the 79% - 86% of the shooting phases of the take-off stage (p < 0.001, F = 9.200, Figure 6). Multiple comparisons of motor primitive 2, also for each type of distance shot, revealed a difference only at 78% - 88% of the shooting action for short distance and long distance (p < 0.001, t* = 4.579, Figure 6). There was no difference in motor primitive 3 for different distance shooting, indicating no difference in the degree of activation of motor module 3 during the shooting action phase for all three distances.
The present study revealed that proficient basketball players consolidate their technical actions into three synergies when executing shooting from short, middle, and long distances, with the number of synergies remaining consistent across distances. According to the results of synergistic decomposition, the three motor modules were activated in chronological order to reach the peak, which could be explained by the muscle synergy characteristic of skilled technical movements (Aoyama et al., 2022). Moreover, the peak activation of each module appeared in different movement phases to correspond to different movements, the peak activation of motor module 1 appeared in the lifting ball stage, the peak activation of motor module 2 appeared in the take-off stage, and the peak activation of motor module 3 appeared in the release stage. And it was discovered that there were some differences between the motor modules and motor primitives between different distance shooting, verifying our hypothesis. Different distances do not seem to affect the number of basketball shooting muscle synergies, but instead, the patterns of muscle synergies. This finding is consistent with a previous study (Hajiloo et al., 2020) on lower limb muscle synergies at different running speeds. The study suggests that increasing running speed, indicative of heightened task difficulty, influences the motor modules and motor primitives of muscle synergy. Notably, differences in muscle synergies for shooting at different distances mainly appeared between short distance shooting and long distance shooting. Long distance shooting requires more energy at longer distances and smaller visual targets than short distance shooting, requiring more control to complete the shot (Brancazio, 1981; Miller and Bartlett, 1996). So, long-distance shooting can be a more difficult task than short-distance shooting, leading to a change in the synergy pattern. And this shift in synergistic patterns may be the key to successful shootings for different distances. Motor primitive 1 and motor primitive 2 differed in the 45% - 59% (lifting ball stage) and 78% - 88% (take-off stage) of the shooting maneuver for short and long distance shooting, respectively (Figure 6). Conversely, the similarity between motor module 1 and motor module 2 was high during the shooting process. The cortex chose to activate these two identical spinal cord modules while taking shots at different distances based on incoming information from the outside world but with different temporal activation patterns for these modules (Saito et al., 2018). The more highly weighted muscles in the motor module 1 for both distance shots are BB and LD, whose main roles are elbow flexion, shoulder stabilization, and the peak activation of this module occurs during the lifting ball stage. Previous studies have demonstrated that short distance shooting has a higher stabilizing position and shot position than long distance shooting (Miller and Bartlett, 1996). The closer the player is to the basket, the more defensive pressure is suffered (Csataljay et al., 2013), which requires the player to lift the ball faster to reach a certain height. This explains why motor module 1 for short distance shooting has a high activation in the lifting ball stage of the shooting action. Motor module 2, conversely, is weighted more heavily on the core and lower extremity muscles, with the primary roles being core stabilization and hip extension versus knee extension, and the peak activation of this module occurring during the take-off stage. Basketball textbooks recommend paying attention to lower limb power and using the lower extremity more when shooting from a distance (Filippi, 2011). Meanwhile, other studies have shown that the energy generated in the lower limb increases as the shooting distance increases, causing the energy transferred to the upper limb to increase (Nakano et al., 2020). However, some studies observed no relationship between lower extremity strength and successful shooting from a distance (Cabarkapa et al., 2022), which may be due to the fact that the ability to energy generated in the lower limbs is not directly applicable to upper extremity shooting performance. Hence, shooting from a distance requires increased activation of the lower extremity muscles to generate more force and an increase in activation of the core muscles to make the torso more stable and the transfer of energy more efficiently. Figure 6 shows that the peak intensity of activation of this module increased and shifted backward with distance, suggesting that a later and stronger activation of the motor module 2 during the take-off stage may be required for long distance shooting. A study of muscle synergies between high-level breaststroke swimmers and beginners in technical movements showed that the timing of activation of synergistic modules differed between high-level breaststroke swimmers and beginners, and suggested that this may be a determining factor in breaststroke swimmers performance (Matsuura et al., 2020). So later and stronger hip and knee extension may also be a factor in the successful completion of long distance shooting. The motor primitive 3 of short and long distance shooting exhibited no difference throughout the shooting phase and peaked in the final release stage. However, there was less similarity between the motor module 3. Changes in muscle weights in the motor modules are a form of adaptation of the athlete's to the external environment (Cheung et al., 2009). In the motor module 3 of short distance shooting, the muscle weights were primarily dominated by upper limb muscles responsible for shoulder flexion, elbow extension, and wrist flexion. Conversely, in long distance shooting, the weights of lower limb muscles such as the GL, RF, and core muscles like the EOM increased compared to short distance shooting. These additional muscles play key roles in knee and ankle extension, core stabilization, and twisting. This suggests that long distance shooting necessitates activation of both upper and lower extremity as well as core muscles during the release stage of the shooting phase. Moreover, it has been proposed that some of the energy generated from the jump can be utilized to optimize ball release during the release stage of long distance shooting (Knudson, 1993). Increased activation of lower limb muscles alongside upper limb muscles during ball release may aim to maximize the utilization of energy generated from the jump. Additionally, research suggests that long distance shooting may require greater utilization of core twisting forces during the release stage. Furthermore, it has been suggested that distal joints may compensate for variability in proximal joints during the release stage of free throw shots (Button et al., 2003), akin to the predominantly upper limb activation observed during the release stage of short distance shooting in the present study. However, with increased distance, the energy demand for the shot rises, and reliance solely on distal joint compensation may be insufficient. Therefore, long distance shooting may necessitate heightened activation of lower limbs and core muscles during the ball release phase compared to short distance shooting. Additionally, synergistic force generation from both the upper and lower limbs, along with the core, should be taken into account. Athletic training may encourage the development of new muscle synergies to accomplish new tasks, change the weighting of existing muscle motor modules, or change the activation pattern of the motor modules in the timing sequence (Sawers et al., 2015). This principle can also be applied to shooting practice. For novice players, mimicking the muscle synergies of upper and lower limbs, as well as core muscles, observed in elite athletes can aid in acquiring new synergies and mastering shooting techniques. For basketball players proficient in short-distance shooting but lacking proficiency in long-distance shooting, adjusting the timing and intensity of lower limb activation during shooting, and leveraging the energy from jumps and body twists during ball release, can facilitate mastery of long-range shooting. Overall, in long-distance shooting, emphasis should be placed on enhancing lower limb strength and core stability, with particular attention to the coordinated activation timing of upper and lower limb, and core muscles. Some limitations in this study include equipment limitations that allowed EMG data to be collected from only nine muscles, and the number of muscles affects the extraction of synergies (Steele et al., 2013). Additionally, in the shooting tests, subjects were instructed to shoot the ball in their most comfortable manner, without considering potential interfering factors such as defensive pressure. Furthermore, the selection of athletes with minimal height differences (primarily backcourt players) aimed to ensure proficiency in shooting (high shooting percentage) and minimize the influence of height and basket height variations on shooting movements. However, this may limit the generalizability of findings to basketball players in other positions. Future research could explore muscle synergies in shooting among players in different positions. Differences in shooting distance do not affect the number of muscle synergies in the shooting maneuver, but, do affect the pattern of muscle synergy. Athletes undergoing training or coaches providing instruction for distance shooting should focus on lower limb force generation. Additionally, attention should be given to the timing of muscle activation in the upper and lower limbs, as well as the core, and the synergistic coordination of these various muscle groups.
| ACKNOWLEDGEMENTS |
The measurement complied with the current laws of the country in which they were performed. The authors have no conflicts of interest to declare. The datasets generated and analyzed during the current study are not publicly available, but are available from the corresponding author who was an organizer of the study. |
|
| AUTHOR BIOGRAPHY |
|
 |
Penglei Fan |
| Employment: PhD Candidate, Department of Physical Education, Jeonbuk National University |
| Degree: MS |
| Research interests: Sports Biomechanics, Neuromuscular Control Strategies, Physical and athletic performance |
| E-mail: fanpenglei@jbnu.ac.kr |
| |
 |
Zhitao Yang |
| Employment: Lecturer, Department of Mechanical and Electrical Engineering, Zhoukou Normal University |
| Degree: PhD |
| Research interests: Sports Biomechanics, Sports training, Performance analysis |
| E-mail: yangzhitao9928@163.com |
| |
 |
Ting Wang |
| Employment: PhD candidate, Department of Physical Education, Jeonbuk National University |
| Degree: MS |
| Research interests: Sports Biomechanics, Sports injury prevention, athletic performance |
| E-mail: wangting@jbnu.ac.kr |
| |
 |
Jiaying Li |
| Employment: Department of Physical Education, Jeonbuk National University |
| Degree: PhD |
| Research interests: Sport and Exercise Biomechanics, Athletic training |
| E-mail: ky277500122@163.com |
| |
 |
Youngsuk Kim |
| Employment: Lecturer, Department of Physical Education Jeonbuk National University |
| Degree: PhD |
| Research interests: Athletic performance, Sports medicine, Sports biomechanics |
| E-mail: Laity456321@gmail.com |
| |
 |
Sukwon Kim |
| Employment: JBNU Professor, Department of Physical Education Jeonbuk National University |
| Degree: PhD |
| Research interests: Sports performance analysis, Athlete training, Gait, Sports biomechanics |
| E-mail: rockwall@jbnu.ac.kr |
| |
|
| |
| REFERENCES |
 Aoyama T., Ae K., Kohno Y. (2022) Interindividual differences in upper limb muscle synergies during baseball throwing motion in male college baseball players. Journal of Biomechanics 145, 111384. Crossref |
Barroso F.O., Torricelli D., Moreno J.C., Taylor J., Gomez-Soriano J., Bravo-Esteban E., Piazza S., Santos C., Pons J.L. (2014) Shared muscle synergies in human walking and cycling. Journal of Neurophysiology 112, 1984-1998. Crossref |
Brancazio P.J. (1981) Physics of basketball. American Journal of Physics 49, 356-365. Crossref |
Button C., Macleod M., Sanders R., Coleman S. (2003) Examining movement variability in the basketball free-throw action at different skill levels. Research Quarterly for Exercise and Sport 74, 257-269. Crossref |
Cabarkapa D., Eserhaut D.A., Fry A.C., Cabarkapa D.V., Philipp N.M., Whiting S.M., Downey G.G. (2022) Relationship between upper and lower body strength and basketball shooting performance. Sports 10, 139. Crossref |
Cabarkapa D., Fry A.C., Cabarkapa D.V., Myers C.A., Jones G.T., Deane M.A. (2021) Kinetic and kinematic characteristics of proficient and non-proficient 2-point and 3-point basketball shooters. Sports 10, 2. Crossref |
Cheung V.C., d'Avella A., Bizzi E. (2009) Adjustments of motor pattern for load compensation via modulated activations of muscle synergies during natural behaviors. Journal of Neurophysiology 101, 1235-1257. Crossref |
Cheung V.C., d'Avella A., Tresch M.C., Bizzi E. (2005) Central and sensory contributions to the activation and organization of muscle synergies during natural motor behaviors. Journal of Neuroscience 25, 6419-6434. Crossref |
Clark D.J., Ting L.H., Zajac F..E., Neptune R.R., Kautz S.A. (2010) Merging of healthy motor modules predicts reduced locomotor performance and muscle coordination complexity post-stroke. Journal of Neurophysiology 103, 844-857. Crossref |
Csataljay G., James N., Hughes M., Dancs H. (2013) Effects of defensive pressure on basketball shooting performance. International Journal of Performance Analysis in Sport 13, 594-601. Crossref |
Csataljay G., O’Donoghue P., Hughes M., Dancs H. (2009) Performance indicators that distinguish winning and losing teams in basketball. International Journal of Performance Analysis in Sport 9, 60-66. Crossref |
Filippi, A. (2011) Shoot like the pros: The road to a successful shooting
technique. Kindle version. T.B. Random House, Chicago.
|
Frère J., Hug F. (2012) Between-subject variability of muscle synergies during a complex motor skill. Frontiers in Computational Neuroscience 6, 99. Crossref |
Hagio S., Kouzaki M. (2015) Action direction of muscle synergies in three-dimensional force space. Frontiers in Bioengineering and Biotechnology 3, 187. Crossref |
Hajiloo B., Anbarian M., Esmaeili H., Mirzapour M. (2020) The effects of fatigue on synergy of selected lower limb muscles during running. Journal of Biomechanics 103, 109692. Crossref |
Hug F., Turpin N.A., Guével A., Dorel S. (2010) Is interindividual variability of EMG patterns in trained cyclists related to different muscle synergies?. Journal of Applied Physiology 108, 1727-1736. Crossref |
Knudson D. (1993) Biomechanics of the basketball jump shot—Six key teaching points. Journal of Physical Education 64, 67-73. Crossref |
Lee, D. and Seung, H.S. (2000) Algorithms for non-negative matrix
factorization. In: Advances in neural information processing
systems. Eds: Dietterich, T.G., Becker, S., and Ghahramani, Z.
Vol. 13. Cambridge, MA: MIT Press. 556-562. |
Li M., Meng X., Guan L., Kim Y., Kim S. (2023) Comparing the Effects of Static Stretching Alone and in Combination with Post-Activation Performance Enhancement on Squat Jump Performance at Different Knee Starting Angles. Journal of Sports Science and Medicine 22, 769. Crossref |
Matsuura Y., Matsunaga N., Iizuka S., Akuzawa H., Kaneoka K. (2020) Muscle synergy of the underwater undulatory swimming in elite male swimmers. Frontiers in Sports and Active Living 2, 62. Crossref |
Miller S., Bartlett R. (1996) The relationship between basketball shooting kinematics, distance and playing position. Journal of Sports Sciences 14, 243-253. Crossref |
Nakano N., Fukashiro S., Yoshioka S. (2020) The effect of increased shooting distance on energy flow in basketball jump shot. Sports Biomechanics 19, 366-381. Crossref |
Norman R.W., Komi P.V. (1979) Electromechanical delay in skeletal muscle under normal movement conditions. Acta Physiologica Scandinavica 106, 241-248. Crossref |
Okazaki V.H., Rodacki A.L., Satern M.N. (2015) A review on the basketball jump shot. Sports Biomechanics 14, 190-205. Crossref |
Okazaki, V.H.A. and Rodacki, A.L.F.J. (2012) Increased distance of
shooting on basketball jump shot. Journal of Sports Science and
Medicine 11, 231. Pubmed |
Pan Z., Liu L., Li X., Ma Y.J. (2023) Characteristics of muscle synergy and anticipatory synergy adjustments strategy when cutting in different angles. Gait and Posture. Crossref |
Park S., Caldwell G.E. (2022) Muscle synergies are modified with improved task performance in skill learning. Human Movement Science 83, 102946. Crossref |
Pataky T.C., Vanrenterghem J., Robinson M.A. (2015) Zero-vs. one-dimensional, parametric vs. non-parametric, and confidence interval vs. hypothesis testing procedures in one-dimensional biomechanical trajectory analysis. Journal of Biomechanics 48, 1277-1285. Crossref |
Safavynia S., Torres-Oviedo G., Ting L. (2011) Muscle synergies: implications for clinical evaluation and rehabilitation of movement. Topics in Spinal Cord Injury Rehabilitation 17, 16-24. Crossref |
Safavynia S.A., Ting L.H. (2012) Task-level feedback can explain temporal recruitment of spatially fixed muscle synergies throughout postural perturbations. Journal of Neurophysiology 107, 159-177. Crossref |
Saito A., Tomita A., Ando R., Watanabe K., Akima H. (2018) Muscle synergies are consistent across level and uphill treadmill running. Scientific Reports 8, 5979. Crossref |
Santuz A., Ekizos A., Janshen L., Baltzopoulos V., Arampatzis A. (2017) On the methodological implications of extracting muscle synergies from human locomotion. International Journal of Neural Systems 27, 1750007. Crossref |
Sawers A., Allen J.L., Ting L.H. (2015) Long-term training modifies the modular structure and organization of walking balance control. Journal of Neurophysiology 114, 3359-3373. Crossref |
Steele K.M., Tresch M.C., Perreault E.J. (2013) The number and choice of muscles impact the results of muscle synergy analyses. Frontiers in Computational Neuroscience 7, 105. Crossref |
Torres-Oviedo G., Ting L.H. (2010) Subject-specific muscle synergies in human balance control are consistent across different biomechanical contexts. Journal of Neurophysiology 103, 3084-3098. Crossref |
|
| |
|
|
|
|