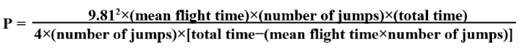|
|
|
| |
| ABSTRACT |
|
Many non-local muscle fatigue studies utilize continuous isometric contractions to induce unilateral fatigue and assess contralateral limb strength and endurance. The present study aimed to investigate the effects of dynamic unilateral lower-limb fatigue on contralateral lower-limb and upper-body explosive performance. Eighteen healthy adults (30.3 ± 9.3 years) completed sessions involving continuous exercise performed to induce fatigue (1 min of unilateral hopping), intermittent exercise performed to induce fatigue (two 30-s bouts of unilateral hopping), and a non-active control. Explosive performance was measured before the intervention and at 1, 5, and 10 min post-intervention using repeated single-leg hops (10 repetitions) and rebound push-ups (5 repetitions). Significant interactions showed that contralateral hop height (p = 0.048), reactive strength index (p = 0.003), and lower-limb power (p < 0.001) were significantly reduced 1 min post-exercise after unilateral continuous hopping. Hop height and power remained significantly lower at 5 min post-exercise (p = 0.03 and p = 0.008, respectively). In contrast, intermittent hopping did not induce significant contralateral lower-limb impairments but resulted in higher upper-body ratings of perceived exertion 1 min post-exercise compared with pre-exercise (p = 0.017) and 5 min post-exercise (p = 0.008). In conclusion, continuous unilateral hopping transiently impairs contralateral lower-limb explosive performance, whereas upper-body explosive performance remained unaffected. Intermittent hopping increases upper-body perceived exertion without affecting contralateral performance. These findings highlight the task-specific and structure-dependent nature of non-local fatigue. |
| Key words:
Crossover fatigue, jump, rebound, reactive strength index, power, contact time
|
Key
Points
- Unilateral dynamic fatigue of the dominant lower limb induced condition-dependent and transient impairments in contralateral lower-limb explosive performance, whereas upper-body explosive performance remained unaffected.
- Continuous loading elicited measurable contralateral decrements, while the incorporation of brief recovery intervals mitigated these effects
- Findings indicate that non-local fatigue is task-specific, centrally mediated, and highly sensitive to the fatiguing stimulus temporal organization (continuous vs. intermittent).
- The rapid recovery supports a predominantly central mechanism and suggests that short recovery periods may be sufficient to restore explosive performance capacity.
|
Neuromuscular fatigue is commonly defined as an exercise-induced reduction in the capacity of the neuromuscular system to generate force or power and reflects the combined influence of peripheral mechanisms within the muscle and central mechanisms within the nervous system (Enoka and Duchateau, 2008; Gandevia, 2001). Although the local or direct consequences of fatigue in the exercised musculature are well established, increasing attention has been directed toward understanding whether fatigue or even stretching can extend beyond the active muscles to influence the performance of muscles that are not directly involved in the task (Behm et al., 2021a; 2021b; Halperin et al., 2015; Miller et al., 2019). Unilateral fatiguing exercise has frequently been used to investigate potential crossover, non-local, or global effects of fatigue. Several studies have reported impairments in non-exercised muscles, most commonly in the contralateral homologous limb (Zijdewind et al., 1998; Rattey et al., 2006; Martin and Rattey, 2007). Because peripheral fatigue mechanisms are confined to the exercised muscle, such crossover effects are generally attributed to centrally mediated processes. These include alterations in supraspinal motor drive and inhibitory feedback from group III and IV muscle afferents (Behm, 2004; Martin and Rattey, 2007; Miller et al., 2019). Despite substantial interest in this phenomenon, findings across studies remain inconsistent. While some investigations have demonstrated reductions in contralateral force production, power, or muscle endurance following unilateral fatigue (Rattey et al., 2006; Martin and Rattey, 2007; Halperin et al., 2014a; 2015), others have observed minimal or no effects, particularly when performance is assessed using brief maximal contractions (Arora et al., 2015; Whitten et al., 2021; Power et al., 2021). These discrepancies suggest that non-local fatigue is not an inevitable consequence of unilateral exercise. One factor contributing to these inconsistencies is the performance task. Evidence from experimental studies and systematic reviews indicates that non-local fatigue effects are more consistently observed during sustained or repeated force production, such as strength-endurance tasks, compared with isolated measures of maximal strength or power (Halperin et al., 2015; Behm et al., 2021a). Furthermore, many non-local fatigue studies intervene with fatiguing isometric contractions (Halperin et al., 2014b; Behm et al., 2021bBehm et al., 2021b,c). However, comparatively little is known about how unilateral dynamic contractile fatigue influences subsequent explosive, multi-joint actions that are highly relevant to sport and functional performance. For this reason, an explosive hop-based task was selected in the present study, as it reflects the rapid force production demands commonly encountered in sport and training settings. In addition to task demands, the structure of the fatiguing exercise itself may influence the expression of non-local fatigue. Exercise performed under sustained conditions has been proposed to promote greater central fatigue and prolonged afferent feedback (Baker et al., 1993; Behm, 2004), potentially increasing the likelihood of non-local performance impairments. In contrast, exercise structures that incorporate brief recovery periods may partially preserve central motor drive (Duchateau et al., 2002) and thus attenuate the accumulation of non-local fatigue. Previous work has suggested that the manner in which fatigue is induced can influence the magnitude of contralateral or non-local effects (Halperin et al., 2015; Behm et al., 2021b; Zahiri et al., 2024). However, few studies have directly examined how different fatigue-inducing exercise structures affect non-local performance outcomes within the same experimental framework. Beyond contralateral lower-limb effects, the potential influence of unilateral lower-limb fatigue on upper-body performance remains poorly understood. Neurophysiological studies have demonstrated that fatiguing exercise performed with the lower limbs can alter corticospinal excitability and intracortical inhibitory and facilitatory processes in upper-limb muscles (Aboodarda et al., 2015; 2017; Šambaher et al., 2016). However, these neural adaptations do not consistently translate into measurable decrements in upper-body performance (Li et al., 2019; Behm et al., 2021b; Whitten et al., 2021). Cross transfer effects from the lower limbs to upper limbs would not only represent contralateral heterologous muscle effects from the same vertebral spinal level but global body influences from the activity of a unilateral leg action. According to Halperin’s et al., review (2015), non-local muscle fatigue effects are more predominant with the testing of the lower rather than the upper body, which could be a manifestation of differences in the neural network of the upper and lower body. Compared to prior published unilateral isometric fatigue protocols, would dynamic fatiguing protocols elicit greater global (lower limb fatigue effects on upper limb performance) responses. Moreover, most studies assessing upper-body responses have relied on isolated strength measures rather than explosive, functionally relevant tasks, limiting the practical interpretation of these findings. Another important limitation of the existing literature is the limited consideration of the temporal profile of non-local fatigue. Many studies assess performance only immediately following the fatiguing task, providing little insight into the persistence or recovery of non-local effects. Emerging evidence suggests that when contralateral impairments occur, they may be transient and resolve within minutes after exercise cessation (Duffett et al., 2021; Zahiri et al., 2024). Systematic assessment across multiple post-fatigue time points is therefore necessary to better characterize the magnitude and duration of non-local fatigue effects. To our knowledge, no study has concurrently examined contralateral lower-limb and upper-body explosive performance following unilateral dynamic fatigue (i.e., hopping) while accounting for both exercise structure and the post-fatigue time course within a single experimental design. Understanding whether unilateral lower-limb fatigue transfers to contralateral or upper-body explosive performance may also have practical implications for the sequencing of unilateral and whole-body exercises within training sessions. Therefore, the purpose of the present study was to examine the effects of unilateral fatigue induced in the dominant lower limb on the performance of the contralateral non-dominant lower limb and upper body during explosive tasks under different experimental conditions. Performance was assessed across multiple post-fatigue time points to characterize the time course of any observed non-local effects. It was hypothesized that fatigue of the dominant leg would result in greater performance impairments in the contralateral, non-dominant, lower limb compared with the upper body, and that any non-local effects observed would be condition-dependent and time-sensitive in nature. ParticipantsAn a priori statistical power analysis was conducted (F test for repeated measures ANOVA within factors) based on prior related research (Halperin et al., 2014bHalperin et al., 2014b, Li et al., 2019, Zahiri et al., 2024) with projected effect size of 0.5, p = 0.05, power = 0.8, number of groups = 3: number of measures = 4. The G *Power (Heinrich Heine University of Dusseldorf, Germany) software predicted that a minimum of 9 participants would be needed to achieve the appropriate power. Hence, to ensure greater than adequate statistical power, 18 healthy adults (13 males, 5 females) voluntarily participated in this study with 17 participants (94.4%) right-arm dominant, and 16 participants (88.9%) right-leg dominant. Arm and leg dominance were assessed as the arm/hand used for writing while leg dominance was assessed as the leg used to kick a ball. Anthropometric characteristics were as follows: Males: age = 28.15 ± 9.30 years; height = 178.77 ± 8.97 cm; body mass = 84.68 ± 11.88 kg; upper-body mass = 57.54 ± 8.97 kg. Females: age = 35.80 ± 7.53 years; height = 164.20 ± 4.32 cm; body mass = 71.58 ± 10.64 kg; upper-body mass = 47.10 ± 7.13 kg. For the upper-body loading mass estimate, a digital scale (Taylor Bath Scale: China) was used. Participants placed their hands on the scale while holding a plank position with a straight body (feet on the floor). The recorded value was used as an estimate of upper-body loading rather than actual mass. All participants completed all experimental sessions. Participants were recreationally active (participated in dynamic exercise more than 2 times per week) and free from known musculoskeletal, neurological, or cardiovascular conditions that could affect performance. Eligibility was confirmed using the Physical Activity Readiness Questionnaire (PAR-Q+) (Bredin et al., 2025). Participants were recruited using convenience sampling through posters, social media announcements, and verbal outreach. Participants were informed not to partake in high volume or high intensity exercise for 24 hours prior to testing, not consume caffeine or nicotine 6 hours prior to testing and were informed to drink fluids (no alcohol) ad libitum 24 hours prior to testing to ensure adequate hydration. Prior to participation, all individuals provided written informed consent, and ethical approval for the study was obtained from the Interdisciplinary Committee on Ethics in Human Research, Memorial University (ICEHR; No. 20251695-HK).
Experimental designParticipants attended the laboratory on four separate occasions (randomized order) for familiarization, control, continuous fatigue, and intermittent fatigue sessions. An initial familiarization session was incorporated to ensure learning effects were minimized. For the randomization, a random number was assigned to each condition for each participant using the random number function in Microsoft Excel and then sorted the values to determine the order of the sessions. Experimental sessions were separated by a minimum of 48 hours to allow for adequate recovery. During the continuous fatigue session, participants completed a single 1-min bout of unilateral hopping using the dominant leg. Based on the findings of Duchateau et al.,(2002) and the Halperin et al., review (Halperin et al., 2015) suggesting that brief recovery periods may partially preserve central motor drive, the intermittent fatigue session, involved two 30-s bouts of unilateral hopping using the dominant leg, separated by a brief rest period (10s). Participants were instructed to maximize the height jumped and minimize the contact time between hops. In the control session, no fatiguing intervention was performed; participants remained seated for a duration matched to the intervention sessions. The intervention session was 3 minutes, (continuous session: start unilateral hopping 1 minute after pre-test, 60 seconds of unilateral hopping and 1 minute until post-testing: intermittent session: unilateral hopping 50 seconds after pre-test, 70 seconds of intervention (2 x 30s with 10s recovery between sets and 1 minute until post-testing). While workload was not directly measured, exercise duration was matched between conditions. Based on the Zahiri et al., (2024) crossover study involving post-intervention measures at 1-, 3- and 5- min, we wished to consider whether dynamic fatigue would have similar or prolonged crossover effects. Hence, performance testing was conducted before the intervention (pre-test) and at three post-intervention time points: 1-, 5-, and 10- min after the intervention. In a prior review (Halperin et al., 2015), we found that the crossover effects were more apparent with dominant limb fatigue and measurement of the non-dominant non-exercised limb. As this study was the first non-local or crossover fatigue study using explosive tasks, we fatigued the dominant limb and observed the non-dominant limb to increase the chances of observing an effect. If significant contralateral changes were found in this study, then subsequent studies could investigate the opposite direction.
General proceduresAt the beginning of each session, participants completed a standardized warm-up consisting of 5 min cycling on a stationary ergometer at a self-selected comfortable pace. Following the warm-up, participants completed pre-test assessments for lower- and upper-body explosive performance. Throughout all fatigue protocols and performance tests, standardized verbal encouragement was provided by the investigator to ensure consistent maximal effort across sessions. Verbal encouragement involved loudly exhorting the word “go” to the participants every 5 seconds during the fatigue protocol and every 2 seconds during the testing protocols. All testing procedures and data collection were conducted by the same investigator.
Performance assessmentsLower-body explosive performance was assessed using a unilateral leg hop test performed with the non-dominant leg. Upper-body explosive performance was assessed using a push-up rebound test. All hop and rebound push-up assessments were conducted using a portable contact mat system (Chronojump Boscosystem; Chronojump, Barcelona, Spain, 59.0 × 84.1 cm). The system records contact and flight times via a pressure-sensitive platform connected to dedicated acquisition software. From these measurements, hop or rebound push-up height (cm), power (Watts), and reactive strength index (RSI; m·s-1) were calculated using established flight-time methods. The Chronojump Boscosystem calculates average mechanical power from flight-time data recorded by the contact mat. The Chronojump Boscosystem provides an indirect estimate of mechanical power derived from jump timing assumptions rather than a direct force-plate measurement. Chronojump's equation for power uses mean flight time and number of jumps as follows:
Based on the actual raw measurements (i.e., flight time), the Chronojump system has demonstrated high validity and reliability for assessing vertical jump performance (ICC = 0.999-1.000; CV = 4.28 ± 1.95%) in active populations (Pueo et al., 2020). However, these estimates should be interpreted cautiously as an indirect proxy derived from flight-time rather than a direct measure of mechanical power. During the push-up rebound test, participants placed their hands directly on the contact mat at shoulder width. With the rebound push-ups, participants had to make contact to the floor with their chest while maintaining a horizontal trunk position. Male participants adopted a standard push-up position (hands and feet in contact with the floor, with a straight trunk), whereas female participants performed a modified push-up with the knees in contact with the floor. Standard push-ups produce a relative (to body mass) ground reaction force of 64-66% whereas modified push-ups with the knees as a contact point exert 49-53% of body mass (Ebben et al., 2011, Gouvali and Boudolos 2005). Based on the reported lesser upper body strength of women (Bartolomei et al., 2021) and our pilot testing, modified push-ups were used for women to allow them to complete multiple repetitions with a rapid ascent phase. For the 5 rebound push-ups, participants were told to “explode” (contract as quickly and forcefully as possible) off the mat and upon landing on their hands to minimize their time on the mat (brief contact time). A similar command was provided for the 10 hops. During the fatiguing hop intervention and hop tests, participants held their hands at the height of the sternum and were allowed to touch the wall with two fingers if they felt unstable when landing. The choice of 5 push-ups and 10 hops was based on pilot work demonstrating that there was greater fatigue with the push-ups and a number of participants could not perform 10 explosive rebound push-ups. The order of lower-body (10 unilateral hops) and upper-body (5 rebound push-ups) performance tests was randomized across participants and sessions to minimize order effects. A 30-second recovery period was allocated between the upper and lower body tests. The investigators observed the participants during testing to ensure proper and standardized technique.
Ratings of Perceived Exertion (RPE)RPE was collected to quantify subjective perceptions of global effort during testing. RPE was recorded using a 10-point category-ratio scale (Borg, 1982), where higher values indicated greater perceived exertion. Participants were familiarized with the scale prior to testing. RPE was verbally obtained and recorded after each pre-test, 1-, 5- and 10- minutes post-tests following the intervention. Ratings were recorded separately after the unilateral leg hop test and the rebound push-up test at each time point.
Statistical analysesStatistical analyses were performed using IBM SPSS Statistics (Version 29.0; IBM Corp., Armonk, NY, USA) and Jamovi (Version 2.5; The Jamovi project). Microsoft Excel (Microsoft 365; Microsoft Corp., Redmond, WA, USA) was utilized for data visualization and the creation of figures. Data in the text and tables are presented as mean ± standard deviation (SD), while data in figures are presented as mean ± standard error of the mean (SEM). The Shapiro-Wilk and Mauchly's Tests were used to assess the normality of the distribution and assumption of sphericity, respectively (P > 0.05). Since the study used a two-way (3 × 4) repeated-measures ANOVA, there were 12 repeated measurements for the RPE variable. In repeated-measures analyses, the statistical model is evaluated based on the assumptions of the repeated-measures ANOVA rather than the distribution of each individual measurement in isolation. For the RPE data, Mauchly's test indicated that the assumption of sphericity was met for the Session, Time, and Session × Time effects. Therefore, the standard repeated-measures ANOVA (sphericity assumed) was appropriate, and no Greenhouse-Geisser correction was required. To determine differences between conditions and time points, a two-way (3 × 4) repeated-measures analysis of variance (ANOVA) was conducted with Session (Intermittent, Continuous, Control) and Time (Pre, 1- min, 5- min, and 10- min after exercise) as within-subject factors. Mean data from the lower body hops (10 repetitions) and upper body rebound push-ups (5 repetitions) were analyzed. Power and RSI were considered secondary outcomes as they were calculated/derived from the primary outcomes of hop or rebound push-up height (calculated from flight time) and contact time. Sex was not included as a factor in the statistical analyses due to the small number of female participants (n = 5), which limited statistical power and precluded meaningful sex-based comparisons. Therefore, analyses were conducted on the pooled sample. When significant main effects or interactions were observed, the Bonferroni adjusted post hoc test was employed to identify specific pairwise differences. Effect sizes were assessed using partial eta squared (ηp2). Statistical significance was accepted at p ≤ 0.05 for all analyses. Partial eta2 values represent effect size magnitudes as follows: 0.01: small, 0.06: medium, 0.14 or higher: large. Reliability of the average hop (ICC (3, 10)) and rebound push-up heights (ICC (3,5)) (height as a representative of all derived measures such as power) was assessed using a two-way mixed-effects, consistency, single-measures intraclass correlation coefficient with fixed rater.
For all tests, the Shapiro-Wilk test of normality was non-significant suggesting data was approximately normally distributed while the Mauchly test of sphericity was also non-significant indicating sphericity was achieved. Reliability as assessed with ICC was considered high with r of 0.97 and 0.86 for hop and rebound push-up height respectively (Table 1). All significant main effect and interaction findings exhibited large magnitude ηp2 effect sizes except for upper body RPE which approached a large magnitude (ηp2: 0.13). As a reminder, hop and rebound push-up height, power, and reactive strength index were derived from their respective flight and contact time values. Non-dominant leg hop performanceDescriptive statistics for non-dominant leg performance are presented in Table 2. There were no significant main effects or interactions for contact time (p > 0.05). However, significant Session × Time interactions were observed for hop height (F(6, 102) = 3.21, p = 0.006, ηp2 = 0.16), RSI (F(6, 102) = 3.54, p = 0.003, ηp2 = 0.17), and power (F(6, 102) = 4.33, p < 0.001, ηp2 = 0.20) (Figure 1). Post hoc Bonferroni-adjusted comparisons revealed that during the continuous session, hop height (p = 0.048), RSI (p = 0.003), and power (p < 0.001) were significantly lower 1 min after exercise compared with pre-exercise values. Additionally, RSI (p = 0.03) and power (p = 0.008) remained significantly suppressed at 5 min post-exercise compared to 1 min post-exercise (Figure 1). Regarding RPE, there was no significant Session × Time interaction (p = 0.97). A significant main effect of Time was observed (F(3, 48) = 6.22, p = 0.001, ηp2 = 0.28). Post hoc comparisons indicated RPE was significantly higher 1 min post-exercise compared with pre-exercise (p = 0.004) (Table 2).
Upper body (rebound push-up) performanceTable 3 differentiates contact times, flight times (raw data with no calculations involved as for example with power which is derived from an equation) and RPE demonstrating that the females modified push-up values demonstrated trivial to small magnitude effect size lower values than men’s standard push-up values. Descriptive statistics for upper-body performance are presented in Table 4. Analysis revealed no significant Session × Time interactions for contact time (p = 0.94), rebound push-up height (p = 0.61), RSI (p = 0.33), or power (p = 0.47). A significant main effect of Time was observed for rebound push-up height (F(3, 51) = 3.16, p = 0.03, ηp2 = 0.16). Post hoc comparisons indicated that rebound push-up height was significantly greater at 10 min post-exercise compared with 1 min post-exercise (p = 0.02). For upper body RPE, a significant Session × Time interaction was observed (F(6, 96) = 2.41, p = 0.03, ηp2 = 0.13). Post hoc analysis revealed that during the intermittent session, RPE was significantly lower at pre-exercise (p = 0.017) and 5 min post-exercise (p = 0.008) compared with 1 min after exercise (Figure 2).
The present study investigated the non-local effects of unilateral dynamic (hopping) fatigue of the dominant lower limb on contralateral, non-dominant, lower-limb hopping and upper-body rebound push-ups explosive performance under continuous and intermittent fatigue protocols. The major findings were that continuous dominant leg unilateral hopping impaired contralateral hop height, RSI, and lower-limb power, whereas non-dominant upper-body explosive performance during the rebound push-up task remained unaffected. Moreover, upper-body RPE (verbally recorded after the hop and rebound push-up tests at 1-, 5- and 10- minutes post-intervention) were higher following intermittent hopping than continuous. Thus, 60 seconds of continuous unilateral, dominant leg, hopping demonstrated greater impairments on contralateral lower-limb explosive performance than on upper-body explosive capacity. Contralateral impairments observed following continuous exercise performed to induce fatigue align with the concept of centrally mediated non-local fatigue (Zijdewind et al., 1998; Rattey et al., 2006; Halperin et al., 2014b). As peripheral fatigue (i.e., metabolic, morphological) is limited to the exercised muscles, reductions in performance of the non-exercised limb might more likely reflect supraspinal and interhemispheric mechanisms. We speculate that sustained unilateral exercise increased group III/IV afferent feedback, which can depress central motor drive and modulate cortical excitability (Aboodarda et al., 2015; 2017; Šambaher et al., 2016). Continuous loading may accumulate inhibitory afferent input, thereby transiently reducing voluntary activation in both the fatigued and homologous contralateral musculature (Halperin et al., 2015; Zahiri et al., 2024). These findings extend previous work by demonstrating non-local effects with dynamic stretch-shortening cycle tasks rather than isolated isometric contractions. The task-specific nature of the observed effects is noteworthy. Explosive jump performance requires more integrated motor control with rapid force development, neuromuscular timing, and coordinated multi-joint activation, possibly making it sensitive to subtle reductions in central drive (Behm et al., 2024). In contrast, prior studies using brief maximal voluntary isometric contractions or single-effort strength tests often reported null contralateral effects, possibly because such tasks insufficiently challenge central mechanisms (Behm et al., 2021b). The current results suggest that dynamic, high-velocity tasks might be more likely than isometric contractions to reveal significant non-local (contralateral, non-dominant, homologous) muscle fatigue (Halperin et al., 2015; Zahiri et al., 2024). The contrast between continuous and intermittent protocols highlights the role of temporal exercise structure in non-local fatigue. Intermittent unilateral hopping, incorporating two 30-second bouts with rest, did not impair contralateral performance, suggesting that brief recovery intervals may help to restore central motor drive, facilitate metabolite clearance, and attenuate afferent-mediated inhibition. This structure-dependent response may explain inconsistencies across previous literature, where sustained protocols frequently elicited contralateral impairments while intermittent or recovery-based designs often did not (Behm et al., 2021b; Zahiri et al., 2024). Despite prior evidence that unilateral lower-limb fatigue can modulate corticospinal excitability in upper-limb muscles (Šambaher et al., 2016; Aboodarda et al., 2017), upper-body explosive performance remained unaffected in this study. These findings may suggest that heterologous muscle groups may be less sensitive to transient central perturbations, particularly during brief explosive tasks. Stronger neural coupling between homologous muscles might explain why contralateral lower-limb effects were detected, whereas anatomically distant upper-body muscles required greater magnitude or duration of central fatigue to show functional deficits. The time course of recovery provides further mechanistic hypotheses. Contralateral impairments were evident 1 min post-exercise but resolved within minutes, suggesting the possibility of a predominantly central origin rather than structural or metabolic changes in non-exercised limbs. As afferent feedback decreases and cortical excitability normalize (Aboodarda et al., 2015, 2017, Šambaher et al., 2016), voluntary activation appears to recover rapidly (Zahiri et al., 2024). Inclusion of multiple post-fatigue assessments in the present study allowed a more nuanced characterization of the transient nature of non-local fatigue. From an applied perspective, these findings indicate that continuous unilateral plyometric exercise may transiently impair contralateral lower-limb explosive performance, whereas brief recovery intervals can mitigate such effects. Practitioners should consider exercise sequencing when unilateral fatigue-inducing tasks precede contralateral explosive movements. The rapid recovery observed suggests that short post-exercise rest may be sufficient to restore performance. LimitationsSeveral limitations should be acknowledged. The sample size was modest, and participants had recreational training backgrounds, limiting generalizability. It is possible that a trained or sedentary group might respond differently as they could have greater or lesser resistance to fatigue and lower or greater metabolic and neural disruptions associated with the exercise protocol respectively. We did not directly measure the extent of the intervention fatigue. However, fatigue was evident from the remarks made by participants after the intervention and visual inspection of their performance indicating poorer hop performance (height, speed, and contact time). Although global RPE was recorded after the testing protocol, it was significantly higher at 1-minute post-test for both the hop and rebound push-up conditions. This significantly higher RPE score at 1-minute may have been influenced by the prior fatiguing intervention as well as the test. We are confident that all participants experienced fatigue from the intervention. Direct neurophysiological measurements were not obtained, precluding definitive conclusions about underlying mechanisms. Unfortunately, we could not recruit a sufficient number of female participants to achieve the statistical power needed to investigate possible sex differences. As the rebound push-up test was modified for the women, the integration of this data for analysis may have introduced some generalizability issues. However, in practical terms, having women perform push-ups with their knees rather than feet as a point of contact is ubiquitous in training and testing as the lower upper body torque allows women with generally absolute and relative lower upper body strength to perform this exercise. Table 3 highlights that sex differences were of trivial to small magnitude between the sexes when comparing standard (males) vs. modified (from the knees for females) rebound push-ups. Future research integrating dynamic fatigue paradigms with measures of voluntary activation, corticospinal excitability, and interhemispheric inhibition would clarify the neural pathways underpinning structure-dependent non-local fatigue.
In conclusion, unilateral dynamic fatigue (hopping) of the dominant lower limb induced condition-dependent and transient impairments in contralateral lower-limb explosive performance, whereas non-dominant upper-body explosive performance remained unaffected. Continuous loading elicited measurable contralateral decrements, while the incorporation of brief recovery intervals mitigated these effects, highlighting the importance of exercise structure in the manifestation of non-local fatigue. Together, these findings indicate that the expression of non-local fatigue is task-specific, may be centrally mediated, and highly sensitive to the temporal organization of the fatiguing stimulus. The rapid recovery observed further supports the possibility of a predominantly central mechanism and suggests that short recovery periods may be sufficient to restore explosive performance capacity.
| ACKNOWLEDGEMENTS |
We thank the participants for their time. The authors report no actual or potential conflicts of interest. While the datasets generated and analyzed in this study are not publicly available, they can be obtained from the corresponding author upon request. All experimental procedures were conducted in compliance with the relevant legal and ethical standards of the country where the study was carried out. The authors declare that no Generative AI or AI-assisted technologies were used in the writing of this manuscript. This research was partially funded through Dr. Behm’s Natural Science and Engineering Research Council of Canada (NSERC) Discovery Grant (RGPIN-223-05861). |
|
| AUTHOR BIOGRAPHY |
|
 |
Mahta Sardroodian |
| Employment: Prof., School of Human Kinetics and Recreation, Memorial University of Newfoundland, St. John’s Newfoundland and Labrador, Canada |
| Degree: PhD |
| Research interests: Neuromuscular responses to exercise and rehabilitation. |
| E-mail: msardroodian@mun.ca |
| |
 |
Hiwa Rahmani |
| Employment: Research Assistant, School of Human Kinetics and Recreation, Memorial University of Newfoundland, St. John’s Newfoundland and Labrador, Canada |
| Degree: PhD |
| Research interests: Integrated neuromuscular and cardiovascular responses to exercise. |
| E-mail: hrahmani@mun.ca |
| |
 |
David G. Behm |
| Employment: Prof., School of Human Kinetics and Recreation, Memorial University of Newfoundland, St. John’s Newfoundland and Labrador, Canada |
| Degree: PhD |
| Research interests: Neuromuscular physiology and sport. |
| E-mail: dbehm@mun.ca |
| |
|
| |
| REFERENCES |
 Šambaher, N., Aboodarda, S.J., Behm, D.G. (2016) Bilateral knee extensor fatigue modulates force and responsiveness of the corticospinal pathway in the non-fatigued, dominant elbow flexors. Frontiers in Human Neuroscience 10, 18. Crossref |
Aboodarda, S. J., Copithorne, D. B., Pearcey, G. E. P., Button, D. C., Power, K. E. (2015) Changes in supraspinal and spinal excitability of the biceps brachii following brief, non-fatiguing submaximal contractions of the elbow flexors in resistance-trained males. Neuroscience Letters 607, 66-71. Crossref |
Aboodarda, S.J., Šambaher, N., Millet, G.Y., Behm, D.G. (2017) Knee extensors neuromuscular fatigue changes the corticospinal pathway excitability in biceps brachii muscle. Neuroscience 340, 477-486. Crossref |
Arora, S., Budden, S., Byrne, J.M., Behm, D.G. (2015) Effect of unilateral knee extensor fatigue on force and balance of the contralateral limb. European Journal of Applied Physiology 115, 2177-2187. Crossref |
Baker, A.J., Kostov, K.G., Miller, R.G., Weiner, M.W. (1993) Slow force recovery after long-duration exercise: metabolic and activation factors in muscle fatigue. Journal of Applied Physiology 74, 2294-2300. Crossref |
Bartolomei, S., Grillone, G., Di Michele, R., Cortesi, M. (2021) A comparison between male and female athletes in relative strength and power performances. Journal of Functional Morphology and Kinesiology 6, 1. Crossref |
Behm, D. G., Alizadeh, S., Drury, B., Granacher, U., Moran, J. (2021b) (2021b) Non-local acute stretching effects on strength performance in healthy young adults. European Journal of Applied Physiology 121, 1517-1529. Crossref |
Behm, D. G., Alizadeh, S., Hadjizadeh Anvar, S., Drury, B., Granacher, U., Moran, J. (2021a) (2021a) Non-local acute passive stretching effects on range of motion in healthy adults: A systematic review with meta-analysis. Sports Medicine 51, 945-959. Crossref |
Behm, D. G., Alizadeh, S., Hadjizedah Anvar, S., Hanlon, C., Ramsay, E., Mahmoud, M. M. I., Whitten, J., Fisher, J. P., Prieske, O., Chaabene, H., Granacher, U., Steele, J. (2021c) (2021c) Non-local muscle fatigue effects on muscle strength, power, and endurance in healthy individuals: A systematic review with meta-analysis. Sports Medicine 51, 1893-1907. Crossref |
Behm, D.G. (2004) Force maintenance with submaximal fatiguing contractions. Canadian Journal of Applied Physiology 29, 274-290. Crossref |
Behm, D.G., Konrad, A., Nakamura, M., Alizadeh, S., Culleton, R., Hadjizadeh Anvar, S., Pearson, L.T., Ramirez-Campillo, R., Sale, D.G. (2024) A narrative review of velocity-based training best practice: The importance of contraction intent vs. movement speed. Applied Physiology, Nutrition, and Metabolism. Crossref |
Borg, G.A.V. (1982) Psychophysical bases of perceived exertion. Medicine & Science in Sports & Exercise 14, 377-381. Crossref |
Bredin, S., Jamnik, V., Gledhill, N., Warburton, D. (2025) The PAR-Q+ and ePARmed-X+. The Health & Fitness Journal of Canada 18, 75-79. |
Duchateau, J., Balestra, C., Carpentier, A., Hainaut, K. (2002) Reflex regulation during sustained and intermittent submaximal contractions in humans. Journal of Physiology 541, 959-967. Crossref |
Duffett, C. A., Wiseman, S. A., Punjabi, A., Power, K. E., Behm, D. G. (2021) Contralateral muscle fatigue from slow, isokinetic contractions is not velocity-specific. The Journal of Sport and Exercise Science. |
Ebben, W. P., Wurm, B., VanderZanden, T. L., Spadavecchia, M. L., Durocher, J. J., Bickham, C. T., Petushek, E. J. (2011) Kinetic analysis of several variations of push-ups. Journal of Strength and Conditioning Research 25, 2891-2894. Crossref |
Gandevia, S.C. (2001) Spinal and supraspinal factors in human muscle fatigue. Physiological Reviews 81, 1725-1789. Crossref |
Gouvali, M.K., Boudolos, K. (2005) Dynamic and electromyographical analysis in variants of push-up exercise. Journal of Strength and Conditioning Research 19, 146-151. Crossref |
Halperin, I., Aboodarda, S.J., Behm, D.G. (2014a) (2014a) Knee extension fatigue attenuates repeated force production of the elbow flexors. European Journal of Sport Science 14, 823-829. Crossref |
Halperin, I., Chapman, D.W., Behm, D.G. (2015) Non-local muscle fatigue: effects and possible mechanisms. European Journal of Applied Physiology 115, 2031-2048. Crossref |
Halperin, I., Copithorne, D., Behm, D.G. (2014b) (2014b) Unilateral isometric muscle fatigue decreases force production and activation of contralateral knee extensors but not elbow flexors. Applied Physiology, Nutrition, and Metabolism 39, 1338-1344. Crossref |
Li, Y., Power, K.E., Marchetti, P.H., Behm, D.G. (2019) The effect of dominant first dorsal interosseous fatigue on the force production of a contralateral homologous and heterologous muscle. Applied Physiology, Nutrition, and Metabolism 44, 704-712. Crossref |
Martin, P.G., Rattey, J. (2007) Central fatigue explains sex differences in muscle fatigue and contralateral cross-over effects of maximal contractions. Pflügers Archiv - European Journal of Physiology 454, 957-969. Crossref |
Miller, W., Kang, M., Jeon, S., Ye, X. (2019) A meta-analysis of non-local heterologous muscle fatigue. Journal of Trainology 8, 9-18. Crossref |
Power, G. M. J., Colwell, E., Hole Saeterbakken, A., Drinkwater, E. J., Behm, D. G. (2021) Lack of evidence for non-local muscle fatigue and performance enhancement in young adults. Journal of Sports Science and Medicine 20, 339-348. Crossref |
Rattey, J., Martin, P. G., Kay, D., Cannon, J., Marino, F. E. (2006) Contralateral muscle fatigue in human quadriceps muscle: evidence for a centrally mediated fatigue response and cross-over effect. Pflügers Archiv - European Journal of Physiology 452, 199-207. Crossref |
Whitten, J. H. D., Hodgson, D. D., Drinkwater, E. J., Prieske, O., Aboodarda, S. J., Behm, D. G. (2021) Unilateral quadriceps fatigue induces greater impairments of ipsilateral versus contralateral elbow flexors and plantar flexors performance in physically active young adults. Journal of Sports Science and Medicine 20, 300-309. Crossref |
Zahiri, A., Goudini, R., Alizadeh, S., Daneshjoo, A., Mahmoud, M. M. I., Konrad, A., Granacher, U., Behm, D. G. (2024) The duration of non-local muscle fatigue effects. Journal of Sports Science and Medicine 23, 425-435. Crossref |
Zijdewind, I., Zwarts, M.J., Kernell, D. (1998) Influence of a voluntary fatigue test on the contralateral homologous muscle in humans? Neuroscience Letters 253, 41-44. Crossref |
|
| |
|
|
|
|